
Abstract
Japanese encephalitis (JE) is one of the most important viral encephalitis, caused by the Japanese encephalitis virus (JEV). The function of non-structural protein 2B (NS2B) mostly remains unclear. In our study, NS2B of Japanese encephalitis virus (JEV) was expressed in Escherichia coli and purified by dialysis. After fusing mouse myeloma cell line SP2/0 with spleen lymphocytes from NS2B protein immunized mice, three clones of monoclonal antibodies (MAbs), named 1B9, 3E12, and 4E6, were generated. The specificity and sensitivity of MAbs were demonstrated by ELISA, indirect immunofluorescence assay, and Western blot. These MAbs will be useful in further exploration of the functions of NS2B and the pathogenesis of Japanese encephalitis virus.
Introduction
J
NS2B is the co-factor of the viral serine protease, which is utilized in stabilization and substrate recognition of NS3 protease. The NS2B-NS3 protease of flaviviruses mediates several cleavages on the flavivirus polyprotein and plays a critical role in the flavivirus replication. Therefore it is believed to be a target for antiviral therapy.(6–8) Substitutions of NS2B-T102M altered JEV replication. JEV NS2B residues Ser46 to Ile60, particularly in Trp53, Glu55, and Arg56, could play an important role in the configuration required for the activity of the flavivirus NS3 protease.(6) JEV infection and the co-expression of JEV NS3 protease with NS2B co-factor induced caspase 3 activation and mitochondria-mediated apoptosis in human medulloblastoma cells.(8) In this study, we used prokaryotic expressed recombinant NS2B as antigen to immunize mice and produced three clones of JEV NS2B-specific monoclonal antibody—1B9, 3E12, and 4E6. The excellent specificity of all the MAbs was demonstrated by indirect immunofluorescent assay (IFA) and Western blotting.
Materials and Methods
Expression and purification of recombinant proteins and mutant proteins
The NS2B gene fragments were amplified from JEV P3 strain (GenBank: U47032) cDNA, which was reverse transcripted from total RNA extraction of JEV-infected BHK cells, using the indicated oligonucleotide primers listed in Table 1. The target fragments were then cloned into the BamHI-XhoI sites of bacterial expression vector pGEX-KG. A full-length glutathione-s-transferase (GST)-NS2B fusion protein expression vector was transformed into Escherichia coli BL21(DE3) and then induced with 1 mmol/L isopropyl-β-thio-galactopyranoside (IPTG) at 37°C for 4 h. Induced expression, bacteria lysis, and purification process of the fusion protein were performed as previously described.(9,10)
The NS2B gene fragments were divided into two parts, including NS2B-1(1-201bp) and NS2B-2(190-393bp). The divided fragments were then cloned into the BamHI-XhoI sites of bacterial expression vector pGEX-KG, respectively. We then performed the mutant proteins using the same method, named GST-NS2B-1 and GST-NS2B-2, with four overlapped amino acids.
Production of mouse monoclonal antibody
Five 4-week-old female BALB/c mice were injected with GST-NS2B protein and Quickantibody adjuvant (Kang Bi Quan Biotech, Beijing, China) into the leg muscle on days 1, 14, and 28, respectively. Three days before fusion, the mice were boosted by intraperitoneal injection with 40 mg of GST-NS2B protein. Splenocytes from immunized mice were harvested and fused to the myeloma cell line SP2/0 with polyethylene glycol 50% (w/v) PEG 4000 (Sigma, St. Louis, MO) at a splenocyte-myeloma cell ratio of 5:1. The fused cells were cultured in HAT medium. Seven days later, the aminopterin was omitted and cells were cultured in HT medium. Hybridoma culture supernatants were screened by ELISA. After subcloning and screening three times, stable antibody-producing clones were left by limiting dilution method and injected intraperitoneally into 8-week-old BALB/c mice. Subsequently, the MAbs were harvested and purified from the seroperitoneum with an antibody purification kit, according to the manufacturer's instructions (NAbTM Protein A/G Spin Kit, Thermo Scientific, Fremont, CA). Their activity was characterized by Western blot analysis and immunofluorescence assay (IFA).
Western blot analysis
The specificity and sensitivity of MAbs were determined by Western blot assay. JEV-infected BHK-21 cell were collected and cell lysate was separated by SDS-PAGE, then transferred overnight at 11 V to nitrocellulose membranes. The membrane was blocked for 1 h with 1% bovine serum albumin (BSA) in TBST buffer (0.01 M Tris-HCl [pH 8.0], 150 mM NaCl, and 0.05% Tween-20), incubated with a 1:1000 dilution of MAbs respectively at room temperature for 2 h. The membranes were reacted with horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG (Southern Biotechnology, Birmingham, AL) for 40 min at room temperature after washing with TBST buffer three times. After repeating the washing step three times, the immune complexes were developed using an enhanced chemiluminescence system.
Identification of MAb subtype
The subtype of the MAbs was identified by a subtype identification kit (Pierce Rapid ELISA Mouse MAb Isotyping Kit, Thermo Scientific, Boston, MA), used according to the manufacturer's instructions.
Indirect enzyme-linked immunosorbent assay
ELISA plates were coated with 0.1 μg/100 μL purified NS2B protein diluted in bicarbonate coating buffer (pH 9.6) at 4°C overnight, blocked with 5% bovine serum albumin (BSA) in PBS for 1 h at 37°C. The wells were drained and incubated with 100 μL/well 2-fold MAb dilutions in PBSA (from 1:2000 to 1:256,000) for 30 min at 37°C. After three washes with PBS containing 0.05% Tween-20 (PBST), 100 μL horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG was incubated for 30 min at 37°C. After washing, 50 μL/well substrate solution A (0.1 M citrate/phosphate buffer [pH 5.0]) and 50 μL/well substrate solution B (0.04% OPD; 0.14% H2O2) were applied for 10 min at room temperature away from light. Reactions were terminated by the addition of 50 μL/well 2 M H2SO4, and optical densities (OD) were measured at 630 nm using a microplate reader. Absorbance values two times higher than the background level reactivity were considered to be positive.
Immunofluorescence assay
BHK-21 cells were cultured in 24-well tissue culture plate and infected with JEV P3 strain. After 36 h post-infection, the cells were fixed with absolute methanol. The MAbs were added in as primary antibody at room temperate for 2 h. After washing three times, cells were overlaid with a 1:500 dilution of fluorescein isocyanate-conjugated goat anti-mouse IgG (Sigma). After 1 h incubation at room temperature, fluorescent images were examined under a fluorescent microscope.
Results and Discussion
Expression of NS2B protein
The recombinant plasmids were transformed into E. coli BL21(DE3) cells and induced with 1 mmol/L IPTG at 37°C for 4 h to express the recombinant protein. As visualized, by SDS-PAGE, the molecular weight of GST-NS2B recombinant protein was 40.4 kDa, consisting of NS2B protein of 14.4 kDa and GST-tag of 26 kDa (Fig. 1).

SDS-PAGE analysis of GST-NS2B protein. Lane 1, protein marker; lane 2, bacilli precipitation of GST-NS2B; lane 3, bacilli precipitation of GST.
NS2B-specific monoclonal antibodies
After fusing mouse myeloma cell line SP2/0 with spleen lymphocytes from NS2B protein immunized mice, three NS2B-specific hybridoma clones (MAbs 1B9, 3E12, and 4E6) were generated and isolated. The stable antibody-producing clones were injected intraperitoneally, and seroperitoneum was harvested for further characterization.
Indirect ELISA and subtype identification
The ELISA titers of 1B9, 3E12, and 4E6 were the same—that is, 1:256,000. The subtype identification result showed that two MAbs of NS2B (3E12 and 4E6) belonged to subtype IgG1 while the other belonged to subtype IgG2b; the light chains of these MAbs were kappa (Table 2).
MAbs specifically recognize NS2B of Japanese encephalitis virus
We used two methods to confirm the specificity of the MAbs. On one hand, BHK-21 cells were infected with JEV P3 strain and then analyzed by immunofluorescence assay. Specific fluorescence signals were observed with MAbs 1B9, 3E12, and 4E6 against NS2B protein. In contrast, the negative control cells were not detected in any fluorescence staining (Fig. 2). On the other hand, Western blot analysis also showed that MAbs 1B9, 3E12, and 4E6 specifically reacted with the protein NS2B of JEV, while no reaction was observed with non-treatment cells (Fig. 3). In conclusion, those clones of MAbs possess high specificity to JEV.
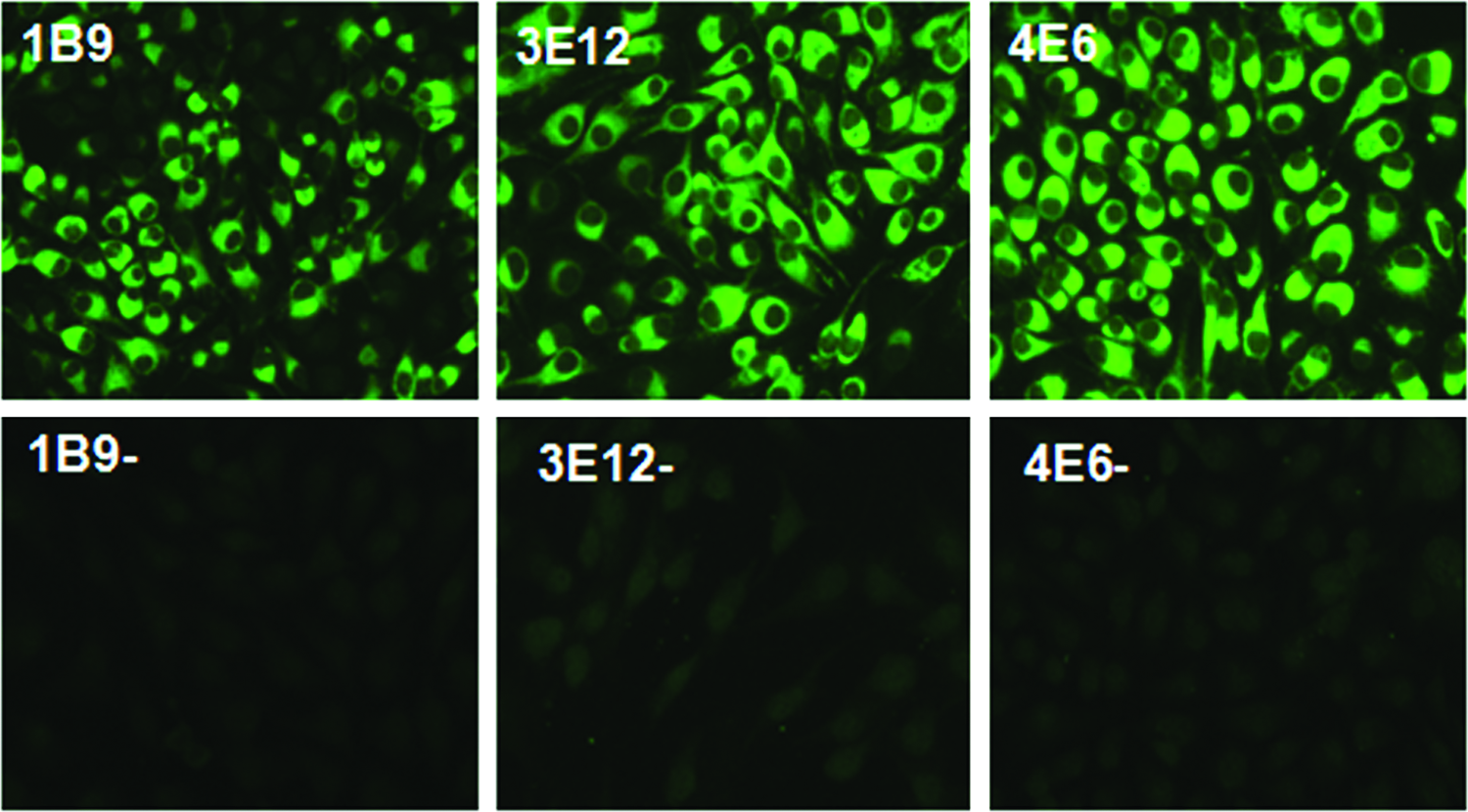
Immunofluorescence staining of JEV P3 strain-infected BHK-21 cells with different MAbs. Fluorescent images were examined with a fluorescence microscope.

Western blot analysis of monoclonal antibodies against NS2B proteins. JEV P3-infected cells were lysed and separated by SDS-PAGE and transferred to nitrocellulose membranes. The membranes were blocked 1 h with blocking buffer, then reacted with a 1:1000 dilution of NS2B-specific MAbs, followed by incubation with HRP-conjugated goat anti-mouse IgG secondary antibody with a 1:3000 dilution. The immune complexes were finally developed by an enhanced chemiluminescence system. The NS2B protein is indicated by arrows on the left side, and its molecular weight is labeled as kilodaltons (kDa). Lanes 1, 3, 5, JEV-infected BHK cell lysate; lanes 2, 4, and 6, BHK cell lysate; lanes 1, 2, incubated with MAb 1B9; lanes 3, 4, incubated with MAb 3E12; lanes 5, 6, incubated with MAb 4E6.
Epitope characterization of MAbs against NS2B
To further narrow the binding region of monoclonal antibody, two mutant proteins with four overlapped amino acids to each other NS2B-1 (1-201bp) and NS2B-2 (190-393bp) were generated. Primer sequences of the expected genes are listed in Table 1. SDS-PAGE results showed that the molecular weight of both mutant proteins were about 33.4 kDa (Fig. 4A), corresponding with the expected molecular weight of fusion mutant proteins GST-NS2B-1 and GST-NS2B-2. ELISA and Western blot results indicated that all clones of MAbs reacted with amino acids from 68 to 131 of NS2B (Table 3 and Fig. 4B). These results may lay the foundation for revealing the structure and functions of NS2B protein.
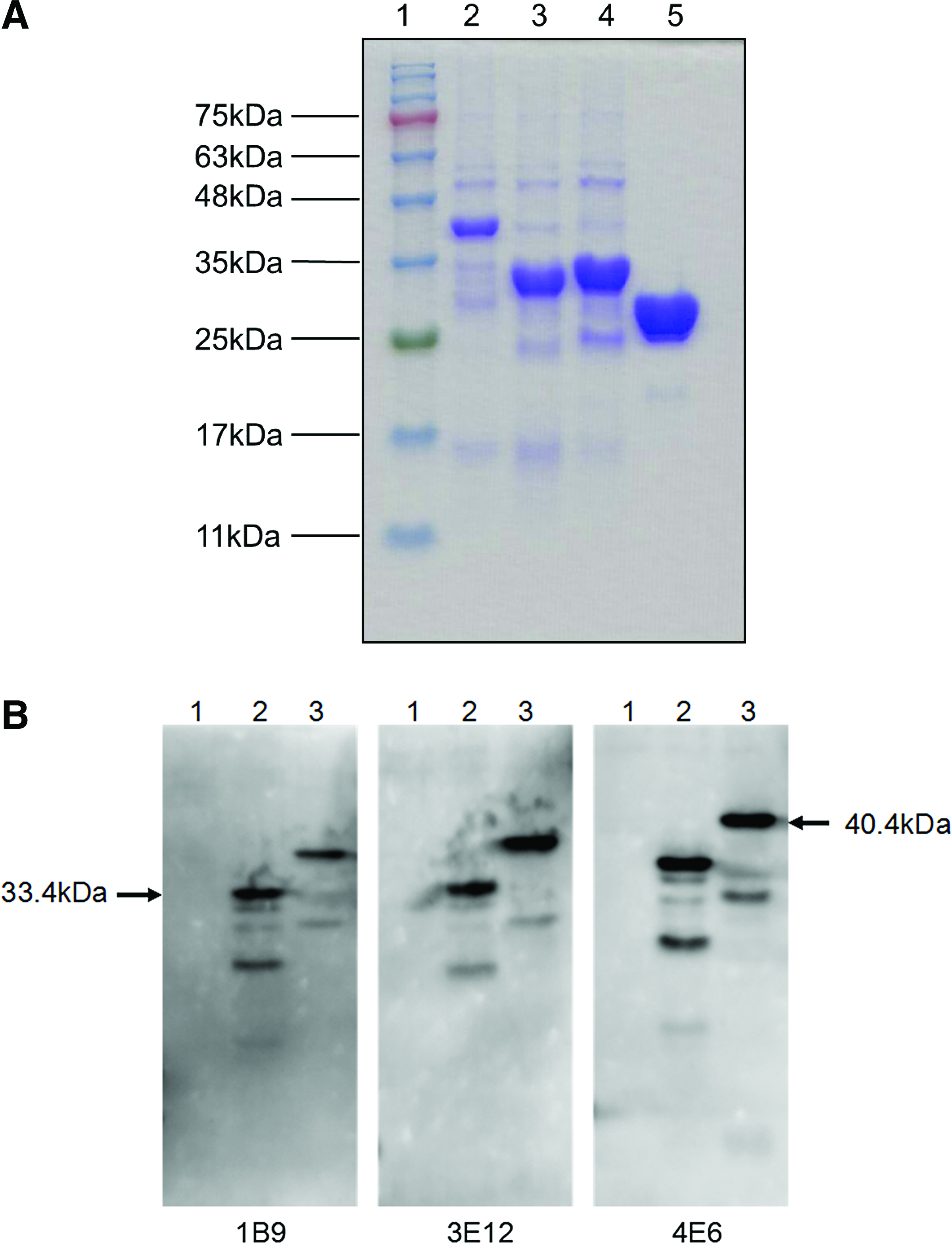
Epitope characterization of MAbs against NS2B. (
Conclusion
Three NS2B-specific monoclonal antibodies were produced and characterized in this study. These MAbs may be used as powerful tools for investigating protein-protein interaction, studying replication mechanism of JEV, and may lay the foundation for future research of cellular and molecular levels of JEV pathogenesis.
Footnotes
Acknowledgments
This work was supported by the National Natural Sciences Foundation of China (31172325), the Special Fund for Agro-scientific Research in the Public Interest (201203082), and the Fundamental Research Funds for the Central University (2011PY002, 2013PY051).
Author Disclosure Statement
The authors have no financial interests to disclose.