
Abstract
During the past few decades, monoclonal antibodies (MAbs) have become an increasingly used tool in diagnostics, therapeutics, and biomedical research. Several methods have been employed to produce MAbs, one of which is the immortalization of B cells by Epstein-Barr virus (EBV). Despite its simplicity, this procedure was never routinely adopted due to its poor efficiency and short-lived antibody (Ab) production. Various adjustments to the basic procedure were introduced, including the addition of certain cytokines and CpG oligodeoxynucleotides, which were shown to improve EBV infectivity and cloning efficiency. The objective of this study was to manipulate culture conditions of the EBV-transformed human lymphocytes, lymphoblastoid cell lines (LCLs), by the timely addition of stimuli including CpG and various interleukins. Such manipulations are aimed at improving LCL proliferative activity and enhancing the cell lines' immortalization potential as well as their Ab production. To accomplish this, IgG+ B cells were isolated from peripheral blood of a hepatitis B vaccinated, anti-HB Ab-positive volunteer. These cells were infected with EBV and incubated in the presence of CpG DNA 2006 motifs, recombinant human interleukin-2 (rhIL-2), rhIL-4, rhIL-6, and rhIL-21, individually and in combinations. Cells were then restimulated for 2 weeks with the same ILs. The effect of these ILs on anti-HB Ab production and the proliferation of the EBV-transformed lymphocytes were investigated. The current study demonstrates that treatment of LCL cultures with rhIL-2, rh-IL4, rhIL-6, and rhIL-21, individually and in combination, increased to varying degrees the proliferative activity and Ab production of these cells. The addition of IL-4 alone was able to sustain increase in anti-HB Ab despite IL-4 withdrawal. This study suggests that with further optimization ILs can have an enhancing effect on LCL immortalization potential and Ab production capacity.
Introduction
I
The production of monoclonal antibodies has progressed significantly over the past 45 years. From the first description of the hybridoma technology by Köhler and Milstein(3 to the advent of chimeric monoclonal antibodies to humanized monoclonal antibodies, successive attempts were made to increase the stability and decrease the immunogenicity of the produced antibodies.(4
Despite generating clinically relevant antibodies, other methods, like phage display, expression cloning, and transgenic mouse approach, all suffered from their own drawbacks and limitations.5,6 Disadvantages of other methods make obtaining human monoclonal antibodies from human peripheral blood B cells a plausible alternative for passive immunotherapy.
Described as early as 1977,7,8 EBV immortalization of B cells uses different sources of human B cells to produce polyclonal B cell activation and proliferation. Having suffered from instability, low clonability, and low affinity of the produced antibodies,5,9 the technique has undergone various modifications to address its shortcomings.
Manipulation of cell culture by the use of interleukins is known to have diverse effects on B cell-transformed cell line activation, immortalization, and/or Ab production. For example, IL-2 is known for its role in stimulating B cell growth and proliferation9,10 while IL-4 increases immunoglobulin secretion(11 without increasing proliferation of LCLs.12,13 IL-6 stimulates growth and immunoglobulin secretion and may improve the stability of these cells(13–15); and IL-21 was shown to promote plasmablast development in an immortalized B cell population, exhibiting enhanced growth and Ab production.15,16
Agonists of Toll receptor 9 such as CpG 2006 were brought to attention when Traggiai and colleagues enhanced the efficiency of EBV transformation by providing innate immune stimulation in combination with EBV treatment to generate Abs against SARS coronavirus.(17 Also, Funaro and colleagues generated potent neutralizing antibodies to CMV by exposing CD22+/IgG+ B cells to CpG2006 and IL-2 and then infecting the cells with EBV.(18
A major mechanism influencing B cell immortalization and stability of Ab production is the BCL-6/Blimp-1 combination. BCL-6 is induced either directly by transducing the cell with constitutionally active STAT5(19 or indirectly using IL-2, IL-4, or IL-21.(15 Effective STAT3-mediated activation of Blimp-1 can also be achieved by a variety of interleukins, including IL-2, IL-6, IL-7, IL-10, IL-15, IL-27, and IL-21.(15
The current study was aimed to develop a procedure to transform B cells by EBV with high Ab production, efficiency, and simplicity by utilizing different combinations of interleukins in the culture medium of EBV-transformed CpG-activated B cells. Such a method can help in further facilitating cost-effective commercial production of therapeutic monoclonal antibodies for various clinical settings.
Materials and Methods
B cell transformation procedure
Peripheral blood from a healthy donor was obtained with informed consent. Peripheral blood mononuclear cells (PBMCs) were isolated from blood by Ficoll hypaque density gradient centrifugation (Sigma-Aldrich, St. Louis, MO). The IgG+ B cells from PBMCs were then isolated using mouse anti-human IgG-coated magnetic microbeads (Miltenyi Biotec, Santa Barbara, CA). PBMCs were incubated with the magnetic microbeads for 15 min, followed by washing with PBS/EDTA buffer and centrifugation. The supernatant was then aspirated completely, and cells were re-suspended in the buffer. Cell suspension was then applied into a magnetic separation (MS) column, which was washed with PBS/EDTA buffer and placed in the magnetic field of a suitable MACS separator (Miltenyi Biotec). Cells labeled with IgG+ microbeads were attached to the column, and the columns were then washed with the buffer three times to eliminate all unlabeled cells that remained unattached. The columns containing the magnetically labeled cells were removed from the separator and placed on a suitable collection tube, and 1 mL of buffer was pipetted onto the column. The labeled cells were then immediately flushed out by firmly pushing the plunger into the column. The magnetic sorting for the isolation of IgG+ cells resulted in a yield of 0.75% (9.5×106 cells out of 1×109 PBMCs).
These cells were immediately used in the B cell–EBV transformation. The IgG+ B cells were initially seeded in U-bottom 24-well plates (Nunc, Roskilde, Denmark) at 0.5×106 cells/well in RPMI 1640 medium supplemented with L-glutamine, 1 mM sodium pyruvate, 50 U/mL penicillin, 50 mg/mL streptomycin, 10% heat-inactivated fetal bovine serum (FBS) (Gibco/Invitrogen Life Technologies, Grand Island, NY), and 10 mM HEPES buffer (PAA Laboratory, Pasching, Austria). All immortalization experiments were conducted in three wells for each tested condition. The isolated B cells were transformed during 2 weeks in the presence of 2×105 autologous irradiated (83 Gy) (Gammacell 1000 Elite Irradiator, Cesium137, MDS Nordion, Kanata, Canada) PBMC feeder cells, 30% v/v EBV-containing supernatant of the B95-8 cell line (ECACC, cat no. 85011419, United Kingdom), 50 U/mL rhIL-2, 100 U/mL rhIL-4, 50 U/mL rhIL-6 (R&D Systems, Minneapolis, MN), 25 ng/mL rhIL-21 (Abcam, Cambridge, MA) and 1 μg/mL CpG2006 (ODN2006, 5′-tcgtcgttttgtcgttttgtcgtt-3′, InvivoGen, Carlsbad, CA). Cell death of PBMC feeder layer was assessed and confirmed at such dose before proceeding with the experiment (data not shown).
Cells were then re-stimulated every 3 days during the first 2 weeks with rhIL-4 (100 U/mL), rhIL-2 (50 U/mL), rhIL-6 (50 U/mL), and rhIL-21 (25 ng/mL) individually and in combination. Cells were then continuously cultured and amplified without further stimulation. Irradiated PBMC feeder layer with CpG2006, EBV, and IgG+ B cells without any cytokine stimulation were used as the control group in the current study.
Anti-HB Ab production and viable cell count
The level of anti-HB Ab production in the supernatant of cells was assessed over a 28-day period. The measurements were performed weekly, using the Architect Anti-HB Ab assay with the chemiluminescent microparticle technique. Also, viable cells were counted weekly by staining an aliquot of the cell suspension with trypan blue (Sigma-Aldrich, Gillingham, United Kingdom). Cells were counted using a Neubauer improved hemocytometer under the light microscope.
Cloning
On day 28, cloning by limiting dilution was performed. The cells that showed high anti-HB Ab production were serially diluted to a final concentration of 1 cell/well and distributed into 96-well plates in the presence of irradiated PBMC feeder layer and interleukins rhIL-2 (50 U/mL), rhIL-4 (100 U/mL), rhIL-6 (50 U/mL), and rhIL-21 (25 ng/mL).
Statistical analysis
All statistical analyses were performed using Microsoft Excel software (2010). The data were analyzed using the one-way analysis of variance (ANOVA) method. A p value of <0.05 was considered statistically significant.
Results
Transformation of B lymphocytes
The magnetic sorting for the isolation of IgG+ cells resulted in a yield of 0.75% (9.5×106 cells out of 1×109 PBMCs). These cells were used for EBV–B cell transformation. After 7 days of EBV infection, proliferating foci of B lymphocytes were seen microscopically. At day 14, the growing foci were macroscopically visible as large clumps of cells. These clumps or aggregates are an indicator that B lymphocytes infected in vitro by EBV were transformed into lymphoblastoid cell lines (LCLs), which are capable of indefinite growth and proliferation (Fig. 1).
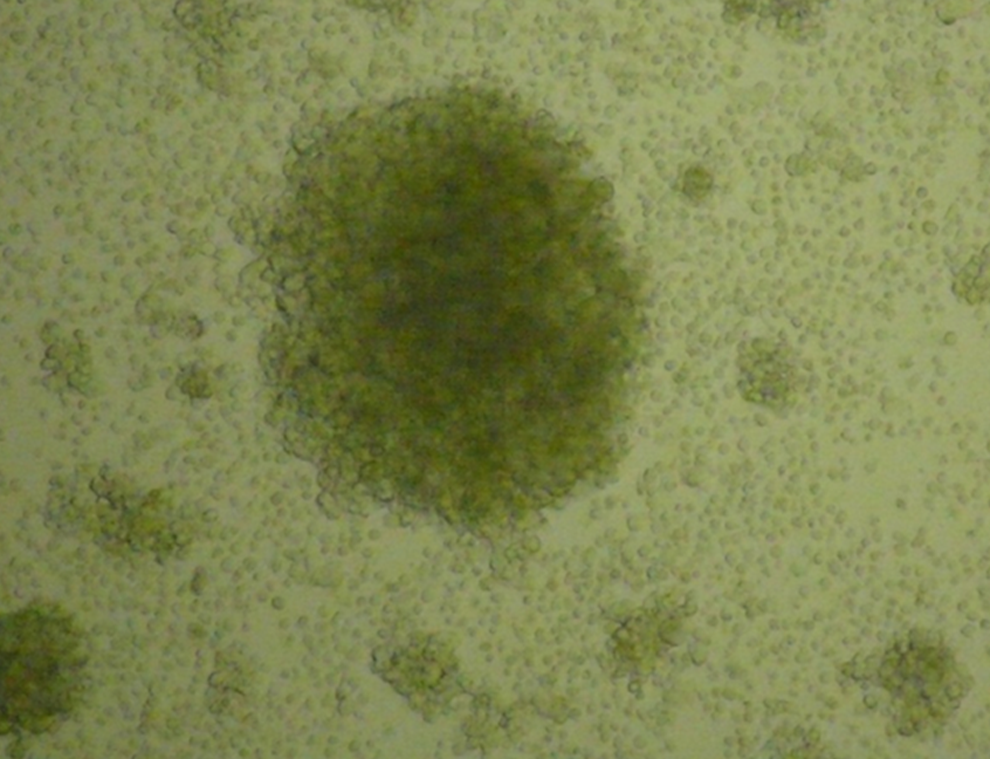
Micrograph showing immortalized B lymphocytes cultured for 2 weeks and treated with rhIL-2 (50 U/mL). 100×magnification.
Effect of various ILs on Ab production of EBV-transformed B cells
Different permutations of interleukins were added to EBV-transformed LCLs; below is a description of the effects of IL-2, IL-4, IL-6, and IL-21 alone and/or in combination on the transformed Ab-producing B cells.
Individual IL treatments
The effect of IL-2, IL-4, IL-6, and IL-21 individually on the proliferation and Ab production by EBV-transformed IgG+ B cells was investigated. The B cells were prepared from a young male positive for anti-HB antibodies after receiving a hepatitis B vaccine booster. EBV-transformed IgG+ B cells were cultured with the following concentrations: rhIL-2 (50 U/mL), rhIL-4 (100 U/mL), rhIL-6 (50 U/mL), and rhIL-21 (25 ng/mL) individually in the presence of CpG DNA 2006 motifs. These ILs were added to each culture every 3 days, starting from the seeding day and for a period of 2 weeks. Anti-HB Ab production of these various treatment groups and the untreated control group were measured weekly for a period of 1 month; the averages of anti-HB Ab levels are shown in Table 1 and Figure 2.

Anti-HB Ab levels produced by LCLs treated with various rhILs over time. The highest production was for cells treated with a combination of rhIL-2 (50 U/mL), rhIL-4 (100 U/mL), rhIL-6 (50 U/mL), and rhIL-21 (25 ng/mL) compared to control cells (irradiated PBMC feeder layer with CpG2006, EBV, and IgG+ B cells). Production peaks on the second week for all interleukins save for IL-4, which has a steady increase in production until week 4.
The following concentrations were used: rhIL-2 (50 U/mL), rhIL-4 (100 U/mL), rhIL-6 (50 U/mL) and rhIL-21 (25 ng/mL) compared to control cells (irradiated PBMC feeder layer with CpG2006, EBV, and IgG+ B cells).
*p=p value.
When comparing the anti-HB Ab production of rhIL-2 treated cells with the control group at all specified time points, it was noticed that these cells had significantly higher production peaking in the second week (13.58±2.38 mLU/mL). This production started to decline in the third and fourth weeks to reach less than 8 mLU/mL. This reduction in the anti-HB Ab levels at both time points could be related to the lack of rhIL-2 in the culture media. A similar pattern was noticed for rhIL-6 and rhIL-2, with a peak on the second week and a decline in Ab production thereafter (Table 1, Fig. 2).
It is worth mentioning that cells treated with rhIL-2 and IL-21 had a slightly higher anti-HB Ab production level in the second week compared to that of IL-6.
Interestingly, cells treated with rhIL-4 showed a different pattern from the other three ILs. These cells showed low levels of anti-HB Ab production at the second week, almost comparable to the control untreated cells, but then showed a significantly higher production of the anti-HB Ab during the third and fourth weeks (Table 1, Fig. 2).
Effect of IL combinations
As for cells treated with a combination of rhIL-2, rhIL-4, rhIL-6, and rhIL-21 compared with the control group at different time points, it was found that this combination had significantly higher anti-HB Ab levels than the control group, with the production peaking during the second week (Table 1). Moreover, although the Ab production declined in the third and fourth weeks, it remained significantly higher than the control, untreated cells as well as most of the individually IL-treated cells.
Effect of various ILs on proliferation of EBV-transformed B cells
Individual IL treatments
The growth stimulatory effect of different recombinant ILs on EBV-transformed IgG+ B cells was tested by counting viable cells after staining with Trypan blue. Apart from rhIL-21 treated cells, all other IL-treated cells showed an earlier increase in growth and proliferation as compared to the control, untreated cells. This earlier increase was most evident in the second week to varying degrees in cells treated with each of rhIL-2, rhIL-4, or rhIL-6 as well as in cells treated with the combination of 4 ILs—rhIL-2, rhIL-4, rhIL-6, and rhIL-21 (Table 2, Fig. 3).

Total cell number of IL-treated LCLs over time. Cell counts of LCL cultures at various time points following treatment with a combination of rhIL-2 (50 U/mL), rhIL-4 (100 U/mL), rhIL-6 (50 U/mL), and rhIL-21 (25 ng/mL) or with rhIL-2 alone were higher than control on all time points. However, cells treated with IL-21 increased in growth after the third week. Control cells (irradiated PBMC feeder layer with CpG2006, EBV, and IgG+ B cells).
The following concentrations were used: rhIL-2 (50 U/mL), rhIL-4 (100 U/mL), rhIL-6 (50 U/mL), and rhIL-21 (25 ng/mL) compared to control cells (irradiated PBMC feeder layer with CpG2006, EBV, and IgG+ B cells).
*p=p value.
The increase in growth and proliferation was most evident for rhIL-2 and the combination of all four ILs, not only in the second week but throughout the entire 4 weeks of the experiment.
As for the rhIL-21 treated cells, although they did not seem to have an early induction of proliferation rate compared to either the other IL treatments or the control, untreated cells, they eventually picked up in their growth rate and reached comparable levels by the fourth week.
Cloning
Cloning by limiting dilution was performed for cells that showed high anti-HB Ab production in this current study. Cloning was done in the presence of an irradiated PBMC feeder layer and interleukins. Successful cloning was noticed in 32 wells out of 160 wells (i.e., cloning efficiency was 20%). However, duplicate measurements of anti-HB Ab in the supernatants of these clones on day 28 showed no Ab production in all clones.
Discussion
Species incompatibility of MAbs produced via the hybridoma technology, in addition to its labor-intensive steps, instigated the development of new or alternative methods for immortalization of B cells. One of these alternative methods utilizes EBV that has been used to immortalize human B cells producing specific antibodies for decades.20,21 However, and despite its simplicity, this method has suffered from low efficiency, lack of reproducibility, and short-term production of antibodies. As in other recent reports, the current study revisits this procedure in an attempt to overcome its shortcomings and optimize the production of MAb from EBV-transformed anti-HB Ab-producing LCLs.
The instability of Ab secretion of lymphoblastoid cells over time could be due to the heterogeneity of the nascent cell line that may contain both Ab-producing LCLs and non-Ab-producing cells. The unwanted non-Ab-producing cells usually have a growth advantage over Ab-producing LCLs. Such heterogeneity could result in domination of the culture by non-Ab-producing LCLs at the expense of the desired producer clones.(22 The instability can also be the due to the upregulation of the activation-induced (DNA-cytosine) deaminase (AID) in LCLs,23,24 which is known to induce somatic hyper mutations (SHM) and isotype switching in B cells.(25 In vivo and in the presence of antigen, SHM leads to increase in Ab repertoire diversity and allows for affinity maturation due to selection of higher affinity antibodies by antigen. However, in vitro and in absence of antigen, this selection does not happen; therefore, SHM could lead to reduction or even loss in Ab affinity during prolonged LCL culture.(26
Our work was focused mainly on adding various ILs alone or in various combinations to enhance both growth properties and Ab production of these LCLs. To this effect, IgG+ B cells were isolated from PBMCs, of HBV immunized subjects, by magnetic selection. Then cells were cultured for 2 weeks in the presence of autologous irradiated feeder cells, 1 μg/mL CpG2006, and EBV containing supernatant. Throughout this 2-week transformed phase, cells were treated with rhIL-2, rhIL-4, rh-IL6, and rhIL-21 individually or in combination. The cultures were then continued without the addition of either stimulus. Control cells (irradiated PBMC feeder layer with CpG2006, EBV, and IgG+ B cells) and IL-treated cells were assessed over a 1 month period for the effect of ILs on the growth of LCLs and anti-HB Ab production. The levels of anti-HB Ab production in the supernatant of irradiated PBMC feeder layer with CpG2006 and EBV were assessed over a 28 day period. All results showed <05.00 mLU/mL, which are considered as negative, according to the anti-HB Ab assay instructions sheet (data not shown). This was done to make sure that anti-HB Abs were produced strictly from LCLs.
Cells treated with IL-2, IL-6, or IL-21 individually and in combination with IL-4 showed the same pattern of anti-HB production where maximum production of Ab was noted in the second week to decline later in subsequent weeks. The decrement in Ab production after the second week could be attributed to the cessation of IL stimulation signals by then.
On the other hand, cells treated with rhIL-4 showed a late but significantly higher production of the anti-HB Abs during the third and fourth weeks, despite the discontinued IL-4 supplementation. This finding agrees with the findings of Shields and colleagues(12 who also found that IL-4 increased production of IgM, IgG, and IgA antibodies from EBV-activated tonsillar B cells. This augmentation was reported to be preferential to low concentrations of IL-4 where 5 U/mL increased Ab production while IL-4 (100 U/mL) did not. However, in another experiment, with cells obtained from a different donor, the LCLs treated with IL-4 (100 U/mL) showed very high Ab production (>15,000 mLU/mL) and continued to produce anti-HB Ab for more than 2 months (data not shown). The differential response to different concentrations of IL-4 may be attributed to IL-4 receptor (IL-4R) polymorphism, where certain IL-4R alleles were reported to induce stronger signals of activation,(27 as well as induction of more IgE production.(28 Moreover, the different effects of ILs depend upon the donors and also on the existence of high- and low-affinity IL-4 receptors coupled to different signaling pathways.(29 The increase in Ab secretion after the second week of the experiment, after cessation of IL-4 additions alone or with the other cytokines, is quite encouraging and could imply a successful immortalization potential and prolonged Ab production when using this cytokine.
As for the effect of interleukins on the proliferation of transformed B cells, the results were fairly consistent for IL treatments apart from IL-21. The most obvious observation was the earlier increase in proliferation as compared to untreated control cells. This was most clear in the second week before stopping the additions of various ILs to the cultures. Such an effect was reported for the some of these ILs, especially IL-2.9,10
However, rhIL-21-treated cells showed lower proliferation rates compared to controls and other ILs, where up to the third week, there was not significant increase in proliferation. However, these cells eventually, by the fourth week, managed to catch up in their growth rate with other cultures. This can be explained in light of a recent study where a differential regulation of B cell proliferation and differentiation by IL-21 was reported. Ozaki and colleagues found that when B cells were treated with IL-21 in vitro, it induced activation of Blimp-1 and Bcl-6, indicating the role of IL-21 as a complex regulator of B cell maturation and terminal differentiation.(16 Also, functional studies demonstrated that IL-21 substantially inhibited proliferation and induced Bim-dependent apoptosis for LPS or CpG DNA-activated B cells.(30 However, the survival in our set may have been promoted due to the presence of EBV transfection as was recently reported in EBV-positive diffuse large B-cell lymphoma cell line where EBV counteracts IL-21-induced apoptosis.(31
Our findings showed that of all the ILs added, singly or in combination, the blend of IL-2, IL-4, IL-6, and IL-21 induced the highest Ab production rates as well as the highest proliferation rates.
Receptors for IL-2, IL-4, IL-6, and IL-21 have been found on the surface of activated B cells.(30,32–34) Thus, it is expected that these ILs acted through their receptors to induce cell division signals, leading to increased proliferation reported in the current study.
As for cloning of producer cells, limiting dilutions down to 1 cell/well have previously enabled researchers to apply the EBV immortalization technology to monoclonal Ab production. When single cell cloning was performed in the presence of irradiated PBMC feeder layer and interleukins, some of the cultures had shown evidence of growth. However, the production of anti-HB Ab was undetectable. This could be reasoned by the unwanted/non-Ab-producing cells, which could have overtaken the culture or they lost their ability to produce more Abs.
Conclusion and Recommendations
The current study has cast a light on the possibility of using EBV-transformed B cells stimulated with proper cytokines to produce sustainable antibody producing cell lines. Future studies would be aimed at including more donors and varying amounts of interleukins due to certain allelic polymorphism of some the cytokine receptors. Also, cells can be treated with various ILs for longer period of time and at different time points during the experiment.
Footnotes
Acknowledgment
Financial support was provided through the Biotechnology Program of the Higher Council of Science and Technology in Amman, Jordan. We thank Dr. Osama Al-Khaldi for his candid support and guidance.
Author Disclosure Statement
The authors have no financial interests to disclose.