
Abstract
G protein-coupled receptor kinase 6 (GRK6) plays a universal role in receptor desensitization, by acting as a receptor-G protein interface, thereby affecting serine/threonine kinases. In this study, a 20-aa-long peptide of human GRK6 C-terminus domain was synthesized and covalently coupled to keyhole limpet hemocyanin (KLH). A mouse monoclonal antibody against human GRK6 (anti-GRK6 MAb) was successfully prepared through hybridoma technique by immunizing BALB/c mice with synthesized GRK6426–446-KLH peptides. A high specificity and affinity strain of hybridoma 5D12 were established. The titer of the purified anti-GRK6 MAb was 1.28×106 measured by indirect ELISA. Western blot and immunocytochemistry experiments were also applied to characterize the antibody specificity. Antibody absorption assays showed that the anti-GRK6 MAb can be blocked by GRK6426–446 peptides. These results indicated that the antibody could bind to GRK6 antigen specifically. This MAb provides valuable support for further studies on the functional properties of GRK6.
Introduction
G
Materials and Methods
Preparation of immunogen
The human GRK6 sequence was subject to a rigorous analysis by using a proprietary algorithm. This method called SEAL™ was designed to identify fragments of protein most likely to yield MAbs. After a careful assessment of peptide scores based on structural features, sequence conservation, hydrophobicity, and solvent, we designed a peptide that is optimized for immunization. The fragment with the highest overall score was selected as the peptide to generate anti-GRK6 MAb. It was then synthesized by Sbsbio (Beijing, China) and subsequently conjugated to KLH using Imject Immunogen EDC Conjugation Kit (Pierce, Rockford, IL) using the manufacturer's protocol.
Animal immunization
The immunization was performed according to methods described previously with minor modifications.(8) During the first immunization, two 8-week-old BALB/c female mice (Dossy Experimental Animals, Chengdu, China) were immunized by intraperitoneal injections with 100 μg GRK6426–446-KLH emulsified by complete Freund's adjuvant (CFA, Sigma, St. Louis, MO). Using the same quantity of antigen with incomplete Freund's adjuvant (IFA, Sigma), two booster immunizations were carried out in 2-week intervals. For the assay of antibody titers, blood from the tail vein was collected after the last immunization. The titers of anti-serum were determined by indirect enzyme-linked immunosorbent assay (ELISA). The mouse with the best immune response was used for antibody development. The final boost without adjuvant was injected intraperitoneally 3 days before cell fusion.
Hybridoma cell production and selection
We followed the cell fusion technique established by Köhler and Milstein.(9) Three days after the final booster injection, the dissociated splenocytes from immunized animals were harvested and fused with myeloma cell line SP2/0 in the presence of 50% polyethylene glycol 4000 (Sigma) at a splenocyte-myeloma cell ratio of 10:1. The fused cells (hybridomas) cultured in 5% CO2 incubator were supplemented with HAT medium (Sigma) containing 10% fetal calf serum, and then distributed in 96-well polystyrene culture plates (Costar, Corning, NY). After incubating for 2 weeks, the culture supernatants producing anti-GRK6 MAb were screened by indirect ELISA using synthesized human GRK6426–446 peptides as coating antigen, and the antibody secreted by the clones were assayed for their binding ability with immunogenic peptides.
ELISA screening
GRK6426–446 (10 μg/mL) in coating buffer was bound to 96-well plates overnight at 4°C. The plates were washed with Tris-buffered saline containing 0.05% Tween-20 (TBST) and then unbound sites were blocked with 2% bovine serum albumin (BSA, Life Technology, Carlsbad, CA) in TBS. After the plates in triplicate were washed with TBST, 60 μL/well of hybridoma supernatants were added into wells and incubated at 37°C for 2 h using culture medium as negative control. After washing three times with TBST, the wells were incubated at 37°C for 1 h in the horseradish peroxidase (HRP)-conjugated anti-mouse IgG antibody (Sigma, 1:1000). After completing the washing steps, 5 mM tetramethylbenzidine (TMB, Vector Labs, Peterborough, United Kingdom) were added. The reaction was stopped with 2 M sulfuric acid after 20 min and the absorbance was measured at 490 nm by a plate reader (Biotek, Winooski, VT). Individual clones were selected by peptides and sub-cloned to yield stable cell lines. The hybridoma cells of positive wells were performed by limited dilution. The highest titer of hybridoma cells was cultured for production of ascitic fluid.
Ascitic fluid production
Two BALB/c mice were intraperitoneally injected with 0.5 mL liquid sterile paraffin oil. Seven days later, each mouse was peritoneally injected with the concentration of 5×106 hybridoma cells/mL producing anti-GRK6 MAb. After 10 days, ascitic fluid was collected and centrifuged at 10,000 g at 4°C for 30 min, and further purified by ammonium sulfate precipitation and rProtein A Sepharose column (Amersham Pharmacia Biotech, Piscataway, NJ) according to the manufacturer's protocol. The purity and concentration of the purified anti-GRK6 MAbs were analyzed by sodium salt-polyacrylamide gel electrophoresis (SDS-PAGE) and Bradford protein assay. Isotype was determined by using mouse monoclonal antibody isotyping reagent kit (Sigma).
Western blot analyses
SH-SY5Y and HT22 cell lines were lysed in RIPA buffer (Millipore, Billerica, MA) supplemented with protease inhibitors (Sigma). Cell lysates were prepared in Laemmli SDS sample buffer (Bio-Rad, Hercules, CA), and protein and were separated by 10% SDS-PAGE. Proteins were transferred to a polyvinylidene difluoride membrane (Millipore) for 1 h at 110 V at 4°C. Membranes were also blocked in TBST containing 5% non-fat dry milk. The 5D12 anti-GRK6 (1:500) primary antibody was used. The appropriate horseradish peroxidase-conjugated secondary antibody was used at 1:10,000 dilution (Santa Cruz Biotechnology, Dallas, TX). Proteins of interest were detected by the enhanced chemiluminescence detection system (Thermo Scientific, Rockford, IL). The size of each band was determined based on the molecular weight of the protein standard (Bio-Rad). In addition, for the antigen absorption experiment, about 100 μg GRK6426–446 protein were incubated with 10 μg of purified anti-GRK6 MAb overnight and immune complexes were used for Western blot analysis.
Immunocytochemistry
SH-SY5Y and HT22 cells were seeded on a 24-well cover glass overnight at 37°C. The culture medium was replaced and cells were fixed with 4% paraformaldehyde for 30 min at room temperature in phosphate buffer (0.1 M). After being washed for 15 min in TBS, samples were blocked in 5% normal bovine serum (Santa Cruz Biotechnology) and 0.5% Triton X-100 for 60 min at room temperature. Cells were incubated with the primary antibody for 24 h at 4°C in TBS containing 1% BSA. After incubation in primary antibody 5D12 (1:200), samples were washed three times for 10 min in TBS. For immunofluorescence staining, Alexa-488-conjugated anti-mouse antibody was used as secondary antibody with DAPI to visualize DNA (Life Technologies, Carlsbad, CA) and incubated for 1 h. After washing in TBS completely, samples were mounted on slides and stored at 4°C. Interest distribution of protein was visualized under a fluorescent laser scanning microscope (Leica, Buffalo Grove, IL).
Results
Immunogen preparation/synthesis of immunogen
A 20-aa-long peptide (KDPAERLGCRGGSAREVKEH) selected from the sequence of C-terminal domain of human GRK6 gene was synthesized and conjugated to KLH using Imject KLH Carrier Protein according to the manufacturer's instructions. GRK6426–446-KLH conjugation was prepared successfully as the immunogen in the generation of specific anti-GRK6 MAb.
Immunization of mice and generation of hybridomas
Two BALB/c mice were immunized with GRK6426–446-KLH protein. The titers of specific antibodies during immunization were monitored by analyzing blood samples using indirect ELISA and displayed high affinity binding with coating antigen. Three days after the final booster injection, the splenocyte fusion experiment was carried out. After a 2-week incubation, the supernatants from hybridoma cells were screened by indirect ELISA. Supernatants of positive wells were subcloned three times by limiting dilution. One stable hybridoma cell line (5D12), which could constitutively produce specific MAb against human GRK6, was successfully established. MAb was then produced in mouse ascitic fluid. The cell line could still stably secrete specific anti-GRK6 MAb after liquid nitrogen freeze-storage. The 5D12 cell line was injected into the abdomen of BALB/c mice for large-scale preparation. After the ascitic fluid was purified, the concentration was 8.353 mg/mL (Fig. 1). The purified anti-GRK6 MAb was then diluted to measure the titers against GRK6426–446 by indirect ELISA. As shown in Figure 2, the purified anti-GRK6 MAb can bind to synthetic immunogenic peptides with a titer of 1.28×106. The heavy chain and light chain of mouse anti-human GRK6 MAb were IgG2a and κ, respectively.

SDS-PAGE analysis of purified anti-GRK6. Lane 1, protein marker; lane 2, purified anti-GRK6 MAb.
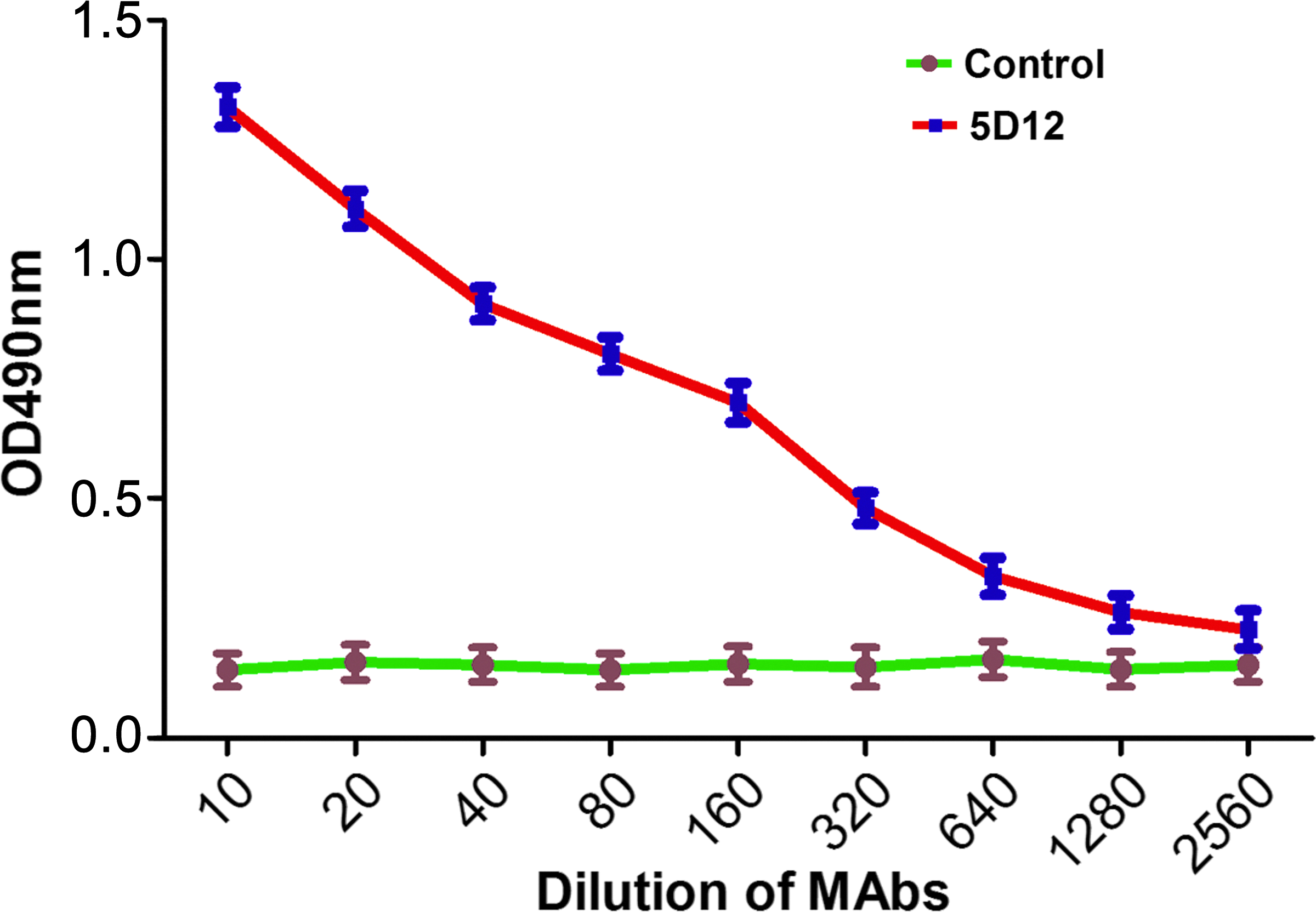
Titer analysis of anti-GRK6 MAb. Line control, SP2/0 mouse ascitic fluid (negative control); line antibody, purified anti-GRK6 MAb.
Western blot analysis
SH-SY5Y and HT22 cells were used to analyze the specificity of anti-GRK6 MAb. The 5D12 MAb can recognize GRK6 protein expressed by the two cell lines. Meanwhile, the Western blot that exhibited a positive band at the molecular weight was not detected after 5D12 MAb was incubated with competitive antigen GRK6426–446 (Fig. 3). The molecular size was about 66kDa using the GRK6 recombinant protein as standard for calibration. This indicates that the antibody generated in our study is specific to GRK6 protein and can recognize natural GRK6 protein. Moreover, non-specific bands were not present in the membrane. According to these results, 5D12 was highly reactive with the cell lysates of different human GRK6. The molecular size was also consistent.
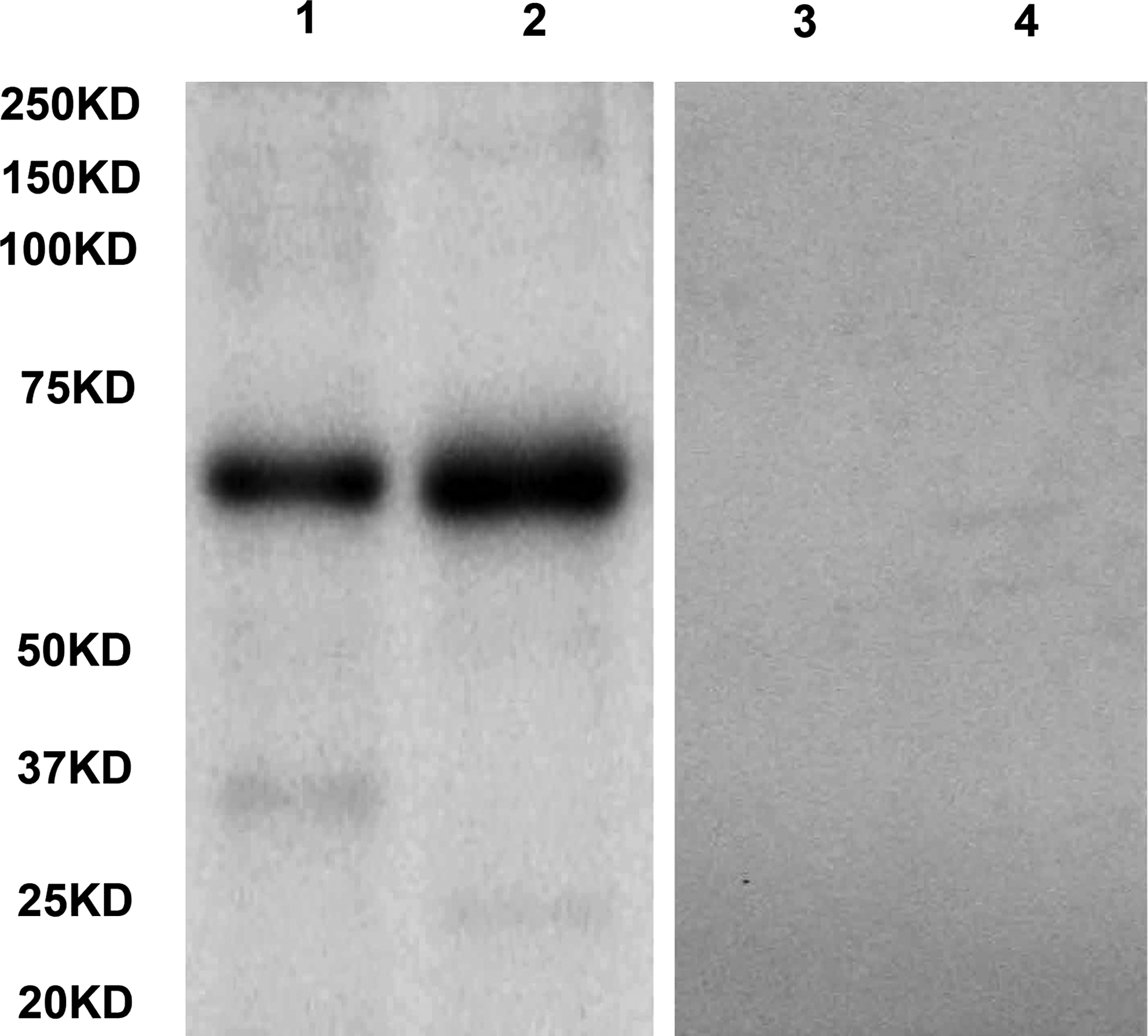
GRK6 was expressed in SH-SY5Y and HT22 cell line. 50 μg of total protein from cell lysates were loaded per lane in SDS-PAGE. Lane 1, 2 SH-SY5Y and HT22 cell lysates were probed with 5D12 GRK6 MAb; lanes 3, 4, SH-SY5Y and HT22 cell lysates were detected with 5D12 antibody after pre-adsorbed with immunizing peptides.
Immunocytochemistry
SH-SY5Y and HT22 cells lines, which can stably express GRK6 protein, were performed for further analysis of the specificity of 5D12 MAb. We found that specific binding signals had the same positive staining directed against GRK6 protein (Fig. 4). The images clearly showed GRK6 protein localized at the cytoplasm and membrane, while by comparison, no signal was detected in the negative control only with the second antibody, thereby proving the specificity of the antibody again.

Specificity of monoclonal antibody analyzed by immunocytochemistry in SH-SY5Y and HT22 cell line (original magnification, x400). Positive signals were shown in 5D12 GRK6 MAb staining.
Discussion
Due to a wide range of tissue distribution, an increasing number of diseases have found a significant correlation with GRK6.(10) Furthermore, the levels of GRK6 are known to change in pathological situations, as seen with heart failure, myocardial infarction, hypertrophy, and hypertension.(3) It plays key physiological and pharmacological roles in the cardiovascular system. Recent studies of GRK6 in the regulation of GPCR substrates have implied critical roles in carcinoma, endocrine diseases, metabolic diseases, gastrointestinal diseases, blood diseases, respiratory diseases, inflammation diseases, musculoskeletal diseases and neurological diseases.(11–17) Therefore, modulation of GRK6 activity and gene expression is an extension of the effort made to discover improved treatments for these diseases. Although a significant amount of investigation is still required to understand the molecular link of GRK6 associated with clinical consequences, GRK6 expression with specialties affords a growing understanding of the biological processes and can be converted into potential diagnostic or therapeutic tools.(18)
Recently, monoclonal antibodies have been considered as a novel immune probe to analyze protein by immunological methods, since they can produce more sensitive and more accurate results in the experiments. In this study, we used a synthetic peptide of human GRK6 protein coupled with carrier protein of KLH eliciting antibody. The mouse anti-human GRK6 MAb was prepared by hybridoma technology. Furthermore one positive hybridoma cell line secreting MAb with high affinity, stability, and specificity was generated and identified. Western blot analysis of human cell lines showed that the MAb obtained can specifically bind to target protein, and the antigen competition experiments of Western blot showed that the anti-GRK6 MAb could be inhibited by GRK6426–446 protein. The specificity of the anti-GRK6 MAb was further proven. The immunocytochemistry result demonstrated that the MAb was specifically directed against human GRK6, comparing the signals of two cell lines. As a valuable tool, the relative specificity of 5D12 provides the basis for the practical study of GRK6 in multiple fields. In the future, we will perform mass spectrometry and high performance liquid chromatography to further define this new antibody.
Footnotes
Acknowledgments
This work was financially supported by the National Natural Science Foundation of China (no. 30671964, People's Republic of China).
Author Disclosure Statement
The authors have no financial interests to disclose.