
Abstract
Folate receptors α (FRα) and β (FRβ) are two isoforms of the cell surface glycoprotein that binds folate. The expression of FRα is rare in normal cells and elevated in cancer cells. Thus, FRα-based tumor-targeted therapy has been a focus area of laboratory research and clinical trials. Recently, it was shown that a significant fraction of tumor-associated macrophages expresses FRβ and that these cells can enhance tumor growth. Although FRα and FRβ share 70% identity in their deduced amino acid sequence, a monoclonal antibody (MAb) reactive with both receptors has not been developed. A MAb that can target both FRα-expressing cancer cells and FRβ-expressing tumor-associated macrophages may provide a more potent therapeutic tool for cancer than individual anti-FRα or anti-FRβ MAbs. In this study, we developed a MAb that recognizes both FRα and FRβ (anti-FRαβ). The anti-FRαβ specifically stained trophoblasts and macrophages from human placenta, synovial macrophages from rheumatoid arthritis patient, liver macrophages from cynomolgus monkey and common marmoset, and cancer cells and tumor-associated macrophages from ovary and lung carcinomas. Surface plasmon resonance showed that the anti-FRαβ bound to soluble forms of the FRα and FRβ proteins with high affinity (KD=6.26×10−9 M and 4.33×10−9 M, respectively). In vitro functional analysis of the anti-FRαβ showed that this MAb mediates complement-dependent cytotoxicity, antibody-dependent cellular cytotoxicity, and antibody-dependent cellular phagocytosis of FRα-expressing and FRβ-expressing cell lines. The anti-FRαβ MAb is a promising therapeutic candidate for cancers in which macrophages promote tumor progression.
Introduction
F
Materials and Methods
Cell lines
B300-19 (murine pre-B cell), KB (human nasopharynx cell), NS1 (murine myeloma cell), RBL2H3 (rat basophilic leukemia cell), and SKOV-3 (human ovarian carcinoma cell) cell lines were obtained from ATCC (Manassas, VA). JFP-J1 hybridoma cells producing anti-GFP rat IgG2a were obtained from Riken BioResource Center (Tsukuba, Japan). The human ovarian carcinoma cell line IGROV-1 was a kind gift from Morphotech (Exton, PA). Both SKOV-3 and IGROV-1 were cultured in RPMI-1640 medium (Life Technologies, Carlsbad CA) supplemented with 10% heat-inactivated fetal bovine serum (FBS), 2 mM L-glutamine, 100 U/mL penicillin, and 100 μg/mL streptomycin (Nacalai Tesque Inc., Kyoto, Japan). The other cell lines were cultured in IMDM medium (Life Technologies) supplemented with 10% heat-inactivated FBS, 2 mM L-glutamine, 100 U/mL penicillin, and 100 μg/mL streptomycin, except for B300-19 cells, which were maintained in the same medium containing 50 μM 2-mercaptoethanol.
Human placenta, rheumatoid arthritis synovium, liver from cynomolgus monkey and common marmoset were obtained from Kagoshima University. Surgical specimens were obtained from 12 patients with ovarian carcinomas and 19 patients with lung cancer at Kagoshima City Hospital and Kagoshima Kousei-Ren Hospital. The diagnoses were made both clinically and pathologically. Tissue samples contained 10 serous and 2 clear cell carcinomas of the ovary, 13 adenocarcinomas, and 6 squamous cell carcinomas of the lung. This study was approved by the ethics committee of Kagoshima University; written informed consent was obtained from all of the subjects.
Production of MAb reactive with FRα and FRβ
FRβ-expressing RBL2H3 and B300-19 were prepared by transfecting human FRβ in the pEF-BOS vector, as previously described.(7,13) Wistar Kyoto rats (Charles River Laboratories, Yokohama, Japan) were injected via the hind footpads with 100 μL of an emulsion containing 1×107 of FRβ-expressing RBL2H3 and Freund's complete adjuvant. After 16 days, the lymphocytes from iliac lymph nodes were fused with NS1 myeloma cells at a ratio of 4:1 in 50% polyethyleneglycol (PEG 1500, Roche, Mannheim, Germany). The hybridoma cells were placed on 96-well plates and cultured IMDM medium supplemented with 10% heat-inactivated FBS and HAT (Life Technologies). After 7 days post-fusion, the hybridoma supernatants were screened by means of immunofluorescence assay against FRβ-B300-19 and KB. One MAb, named clone 5 (rat IgG2a) was selected for its reactivity with both KB and FRβ-B300-19. The hybridoma culture supernatants were purified using goat anti-rat IgG-agarose affinity chromatography (Rockland, Gilbertsville, PA) and used for further experiments. The F(ab’)2 fragment was prepared using pepsin-agarose (Sigma-Aldrich, St. Louis, MO) according to the manufacturer's instructions. All animal studies were performed in accordance with the ethical guidelines for animal experiments at Kagoshima University.
Human peripheral blood mononuclear cells (PBMCs), monocytes, and M-CSF-differentiated macrophages were prepared from the peripheral blood of four healthy donors, as previously described.(7,13) Briefly, PBMCs were isolated using a Ficoll-hypaque density gradient, and monocytes were separated from PBMCs by adhesion to culture dishes. To obtain macrophages, monocytes were cultured in IMDM containing 10% FBS and 25 ng/mL recombinant human M-CSF (PeproTech, Rocky Hill, NJ) for 7 days.
Flow cytometric analysis
After blocking non-specific binding with 10% human-type AB serum, the cells were incubated with 1 μg of anti-human FRα (Mov 18, murine IgG1, which was a gift from Dr. S. Canevari, Istituto Nazionale per lo Studio e la Cura dei Tumori, Milan, Italy); anti-human FRβ (94b, murine IgG1, which was produced in our laboratory)(7,11); clone 5; or one of the following two isotype-matched MAbs: murine IgG1 (MOPC-21, BD Pharmingen, San Jose, CA) or rat IgG2a (R35-95, also from BD Pharmingen) in PBS containing 1% bovine serum albumin and 0.1% NaN3. To examine the effect of soluble FRγ in human serum, KB and FRβ-expressing B300-19 were incubated with clone 5 or rat IgG2a in the presence of 50% of human-type AB serum. After washing with PBS containing 1% bovine serum albumin and 0.1% NaN3, cells were stained with fluorescein isothiocyanate (FITC)-labeled goat F(ab’)2 anti-rat IgG (H+L) (Leino Technologies, St. Louis, MO) or FITC-labeled goat F(ab’)2 anti-mouse IgG (H+L) (Southern Biotech, Birmingham, AL). For dual-color staining of PBMCs and macrophages, cells were incubated with clone 5, FITC-labeled murine anti-human CD3 (UCHT1, Bio-Rad Laboratories, Hercules, CA), Alexa Fluor 488-labeled murine anti-human CD14 (HCD14, BioLegend, San Diego, CA), FITC-labeled murine murine anti-human CD20 (2H7, Bio-Rad), rat IgG2a, or Alexa Fluor 488-labeled isotype control MAbs (BD Pharmingen). The reactivities of clone 5 and rat IgG2a were further detected with APC-labeled goat F(ab’)2 anti-rat IgG (sc-3832, Santa Cruz Biotechnology, Dallas, TX). The stained cells were analyzed using a CyAn™ ADP flow cytometer (Beckman Coulter, Brea, CA).
Immunohistochemical analysis
Immunohistochemical analysis was performed as previously described.(7,14) Briefly, acetone-fixed frozen sections (6 μm) were incubated with anti-FRα, anti-FRβ, clone 5, anti-CD68 (EBM11, Dako, Copenhagen, Denmark) for human tissues, anti-CD68 (Y1/82A, BD Pharmingen) for cynomolgus monkey and common marmoset tissues, or isotype-matched MAbs (murine IgG1, murine IgG2b or rat IgG2a). The staining was visualized with a horseradish peroxidase-conjugated MAX-PO secondary antibody kit (Nichirei Co., Tokyo, Japan) and Nova-Red substrate (Vector Laboratories, Burlingame, CA), according to the manufacturer's instructions. Slides were archived using a Digital Sight CCD camera (DS-Fi1, Nikon, Tokyo, Japan) and computer-aided image analyzer (NIS-Elements, Nikon). The staining intensity was assessed in 10 randomly selected 400x fields in each section by two investigators who were blinded to all clinical details. Positive staining in tumor tissues was defined as staining of >1% of the total cell number.
Production of recombinant soluble FRα and FRβ proteins
Recombinant soluble FRα-Fc protein was prepared as described previously.(15) Briefly, human FRα cDNA (amino acid residues 23-234) without signal peptide and glycophosphatidylinositol anchor peptide was prepared from RT-PCR product derived from KB cells. The primer sequences were 5′-tccaggttccactggtgaccagacaaggattgcatgggccaggactg-3′ (upstream) and 5′-
FRα-Fc protein was purified from culture medium supernatants by Protein A affinity column chromatography (GE Healthcare, Tokyo, Japan). The human FRβ cDNA (amino acid residues 1-230 containing the Kozak consensus sequence) without the glycophosphatidylinositol anchor peptide was amplified from human FRβ in the pEF-BOS vector by the following primer sets: 5′-agaaagacatggtctggaaatggatg-3′ (upstream) and 5′-tcaattcacatgcatggctgcagcatagaacc-3′ (downstream). The PCR product was ligated into the pCR2.1-TOPO vector (Life Technologies), and the KpnI/XhoI-digested insert was then ligated into a pBacPAK9 baculovirus transfer vector (Clontech Laboratories, Mountain View, CA). The recombinant baculovirus was produced by transfecting Sf21 cells (Clontech) with pBacPAK9-FRβ and BacPAK6 viral DNA (Clontech) using the BacPAKTM Baculovirus Expression System (Clontech) according to the manufacturer's instructions. Suspension cultures of Sf21 cells (2×106 cells/mL) were infected with the recombinant virus stock at a multiplicity of infection of 10 for 2 days. FRβ was purified from culture medium supernatants by affinity chromatography using clone 5 immobilized on NHS-activated Sepharose 4 Fast Flow (GE Healthcare). The purity of these proteins was evaluated by 7.5–15% gradient SDS-PAGE.
Surface plasmon resonance
Clone 5 was diluted in 10 mM sodium acetate buffer (pH 4.5) to a final concentration of 18 μg/mL, and coupled to a general layer medium (GLM) chip (Bio-Rad) using a ProteOn Amine Coupling Kit (Bio-Rad). Remaining activated carboxylic groups were quenched with ethanolamine. The immobilization level was approximately 13,000 RU. The blank channel remained empty to serve as a reference. The purified analyte (FRα-Fc or FRβ) was injected into the ProteOn XPR36 system (Bio-Rad) at four different concentrations (25, 12.5, 3.16, and 1.5 μg/mL). Regeneration was performed with 10 mM glycine–HCl (pH 2.5). All binding sensorgrams were collected, processed, and analyzed using the integrated ProteOn Manager software (Bio-Rad). Binding curves were fitted using the Langmuir model describing a 1:1 binding stoichiometry. Each individually captured antibody interacting with the four concentrations of antigen was fitted using ka, kd, and KD. ka is the association rate constant for the antibody–antigen binding, kd is the dissociation rate constant for the pair, and KD is the equilibrium dissociation constant (kd/ka).
Complement-dependent cytotoxicity (CDC) activity
CDC activity was performed as described in the literature with slight modification.(17) Briefly, target cells (KB or FRβ-B300-19)were seeded at 5×104 cells/well in 96-well flat-bottom plates in quadruplicate. In preliminary experiments using concentrations of 0.1, 1, 10, and 25 μg/mL of clone 5, the optimal concentration for CDC was determined to be 10 or 25 μg/mL. One hundred microliters of clone 5 or control rat IgG2a at 10 μg/mL was added to each well and incubated for 15 min at room temperature. Then, 10% baby rabbit complement (CL3441, Cedarlane Laboratories, Ontario, Canada) was added to each well and incubated for 2 h at 37°C and 5% CO2. For spontaneous or maximum controls, target cells without the MAb and complement were added to medium alone or 1% Triton X-100, respectively. The cytotoxicity was evaluated by colorimetric assay using a Cell Counting Kit-8, according to the manufacturer's instructions (Dojindo, Kumamoto, Japan). CDC activity was calculated as cytotoxicity (%)=(experimental colorimetric assay of target cells – spontaneous colorimetric assay of target cells)/(maximum colorimetric assay of target cells – spontaneous colorimetric assay of target cells)×100.
Antibody-dependent cellular cytotoxicity (ADCC) activity
Spleens from Wistar rats were minced, homogenized in IMDM, and centrifuged after passing through a cell strainer (BD Pharmingen). The cell pellet was suspended in RBC lysis buffer (Sigma-Aldrich), and these splenocytes were used as effector cells. Target cells (KB or FRβ-B300-19) were seeded at 5×104 cells/well in 96-well U-bottom plates in quadruplicate. In preliminary experiments using concentrations of 0.1, 1, 10, and 25 μg/mL of clone 5, the optimal concentration for ADCC was 10 or 25 μg/mL. One hundred microliters of clone 5 or rat IgG2a at 10 μg/mL and effector cells (PBMCs or rat splenocytes) at 5×106 cells were added to each well at an effector cell/target cell ratio of 100:1, and further incubated for 4 h at 37°C and 5% CO2. The cytotoxicity was measured by lactate dehydrogenase release in culture supernatants (Cytotoxicity Detection Kit plus, Roche Applied Science, Penzberg, Germany), according to the manufacturer's instructions. ADCC activity was calculated as cytotoxicity (%)=(experimental release of target cells – spontaneous release of effector cells – spontaneous release of target cells)/(maximal release of target cells – spontaneous release of target cells)×100.
Antibody-dependent cellular phagocytosis (ADCP) activity
KB were labeled with CellVue Claret Far Red (Sigma-Aldrich) and seeded at 2×104 cells/well in 96-well U-bottom plates in quadruplicate. In preliminary experiments using concentrations of 0.1, 1, 10, and 25 μg/mL of clone 5, the optimal concentration for ADCP was 10 or 25 μg/mL. One hundred microliters of clone 5, its F(ab’)2 fragment or rat IgG2a at 10 μg/mL, and macrophages (1×105) at an effector cell/target cell ratio of 5:1 were added to each well, and further incubated for 4 h at 37°C and 5% CO2. After a 4-h incubation, cells were harvested and blocked with human-type AB serum, and subsequently stained with a mixture of Alexa Fluor 488-labeled murine anti-human CD11b (ICRF44, BD Pharmingen) and Alexa Fluor 488-labeled murine anti-human CD14 to identify macrophages. The cell-associated fluorescence of Alexa 488/FITC (FL1, macrophages) and Far Red (FL4, target cells) was measured by flow cytometry. ADCP activity was calculated as ratios (%) of KB counts phagocytosed by macrophages to total KB counts.
Statistical analysis
Statistical analyses were performed using the nonparametric Mann–Whitney U-test. A value of p<0.05 was considered to be statistically significant.
Results
Clone 5 MAb reacts with FRα- and FRβ-expressing cells
Clone 5 MAb reacted with the FRα-expressing cell lines IGROV-1, KB, and FRβ-transfected B300-19, but not with FRα-negative SKOV3 and B300-19 (Fig. 1A). In addition, clone 5 reacted with a small fraction of monocytes (CD14+) and with most macrophages cultured with M-CSF, but not T cells (CD3+), B cells (CD20+), and natural killer cells (CD16+) (Fig. 1B). In immunohistochemistry experiments, clone 5 stained FRα-expressing trophoblasts and FRβ-expressing macrophages from human placenta, FRβ-expressing macrophages from rheumatoid arthritis synovium, and liver macrophages from cynomolgus monkey and common marmoset (Fig. 2A, B). In addition, clone 5 stained a large fraction of cancer cells and a significant fraction of TAMs in ovarian and lung carcinomas (Fig. 3). In greater detail, clone 5 stained cancer cells and/or macrophages in six out of 10 samples of serous carcinoma of the ovary (60%) and six out of 13 samples of adenocarcinomas of the lung (46%), but not in clear cell carcinoma of the ovary and squamous cell carcinoma of the lung (Table 1).
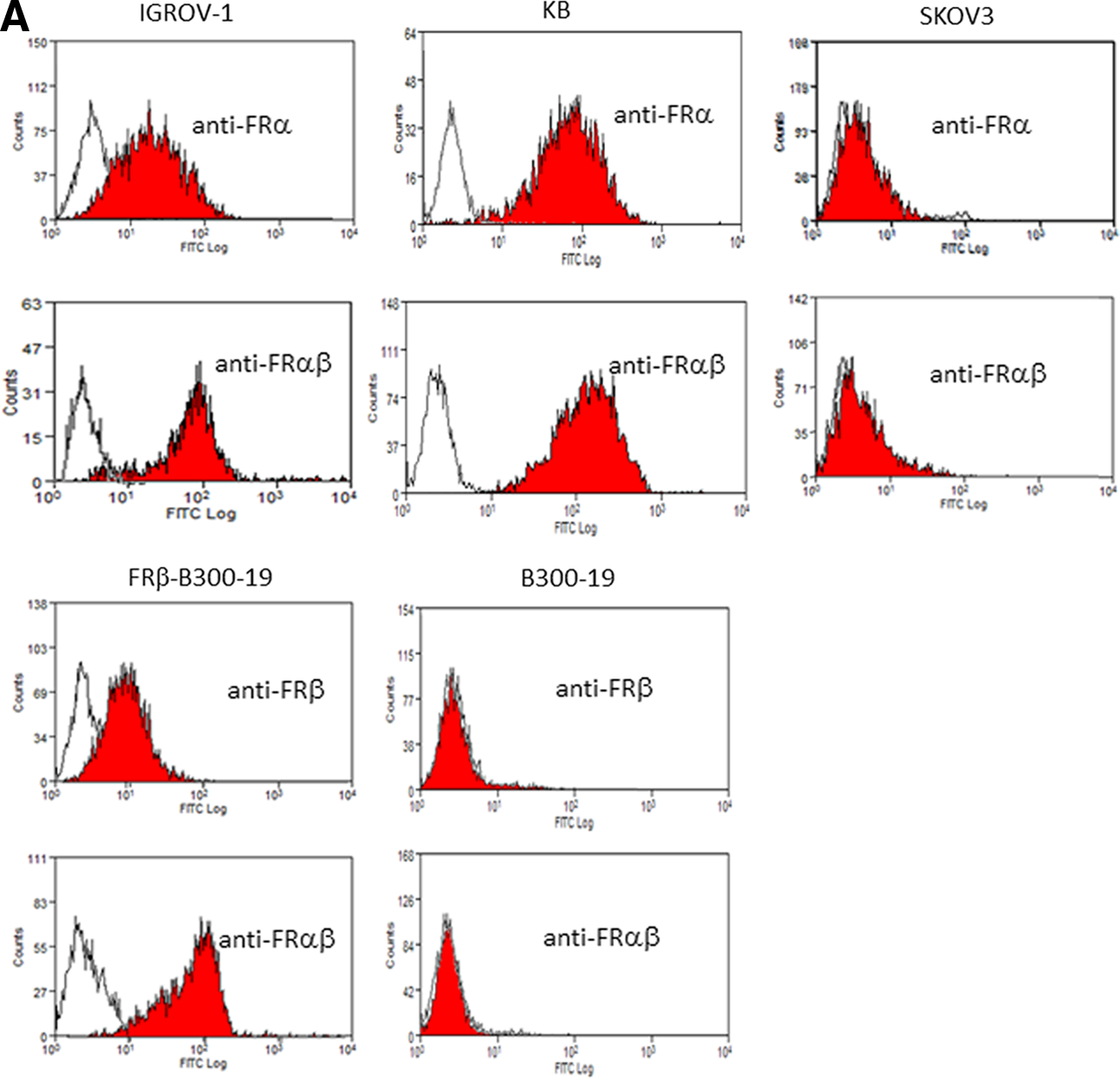
Reactivity of clone 5 with FRα-expressing and FRβ-expressing cells. (

Presence of FR-expressing cells in placenta, rheumatoid arthritis synovium, and liver from cynomolgus monkey and common marmoset. (

Presence of FR-expressing cells in carcinomas. Ovarian and lung carcinomas were incubated with anti-FRα (Mov18), anti-FRβ (94b), anti-FRαβ (clone 5), anti-CD68 (EBM11), or isotype-matched control MAb (mouse IgG1 or rat IgG2a), and the staining patterns were evaluated by immunohistochemistry as described in Materials and Methods. This figure is representative of six positively stained serous lung carcinomas and six positively stained lung adenocarcinomas. Original magnification, 200×.
The previous report suggested the presence of soluble FRγ in human serum.(4) We next examined whether human serum inhibits the binding of clone 5 to FRα- or FRβ-expressing cells in order to know the reactivity of clone 5 with FRγ. As shown in Figure 4, the binding was not inhibited in the presence of human serum.
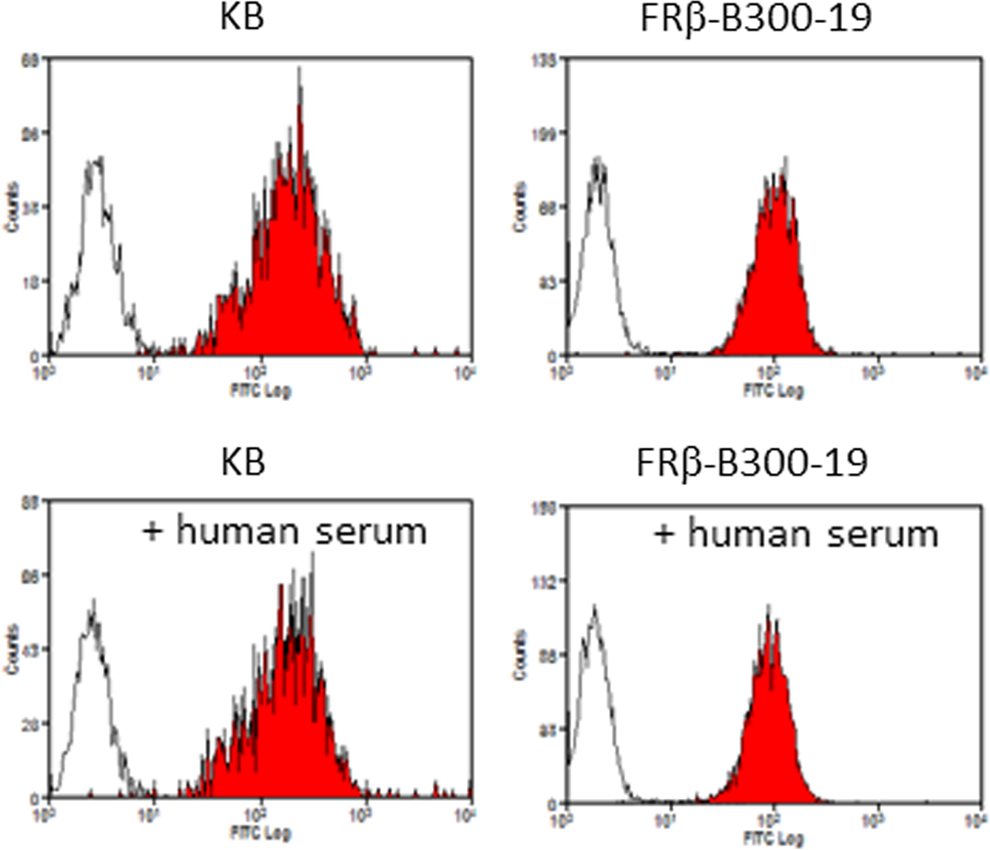
Effect of human serum on reactivity of clone 5 with FRα-expressing and FRβ-expressing cells. FRα-expressing KB and FRβ-transfected B300-19 were incubated with clone 5 or isotype-matched rat IgG2a in the presence or absence of human-type AB serum. The cells were analyzed by flow cytometry as described in Materials and Methods. White and hatched histograms show fluorescence signals with the IgG2a and clone 5, respectively.
Clone 5 MAb has high affinity for soluble FRα and FRβ
Soluble FRα, containing the human IgG1-Fc portion, and soluble FRβ were produced as described in Materials and Methods. The purified soluble FRα and FRβ proteins migrated as single bands of 65 kDa and 22 kDa, respectively, in SDS-PAGE, consistent with their estimated molecular weights (Fig. 5A). In the surface plasmon resonance analysis, soluble FRα-Fc and FRβ reacted with immobilized clone 5 on sensor chips with high affinity (KD=6.26×10−9 M and 4.33×10−9 M, respectively) (Fig. 5B).

Determination of apparent steady-state dissociation constants for the interaction of clone 5 with FRα and FRβ. (
In vitro functional analysis of clone 5 with FRα- and FRβ-expressing cell lines
We examined whether clone 5 mediates CDC and ADCC in FRα-expressing KB and FRβ-transfected B300-19. Clone 5 significantly mediated CDC in these cells in the presence of rabbit complement relative to the control MAb (Fig. 6A). A previous report indicated that rat IgG2a could mediate ADCC by human natural killer cells.(18) Clone 5 MAb mediated considerable ADCC in FRα-expressing KB and FRβ-transfected B300-19 by human blood mononuclear cells; however, this level of ADCC was not statistically significant compared to that mediated by the control MAb. In contrast, clone 5 significantly mediated ADCC in FRα-expressing and FRβ-expressing cells by rat spleen cells compared to the control MAb (Fig. 6B). Next, ADCP was examined using FRα-expressing KB and macrophages. In Figure 7A, regions 1 and 4 show Far Red-labeled KB cells and macrophages stained with the mixture of Alexa 488-labeled anti-CD11b and anti-CD14 MAbs, respectively, while KB phagocytosed by macrophages are shown as double-positive cells in region 3. Clone 5 significantly mediated the phagocytosis of KB cells by macrophages compared to the control MAb, whereas the F(ab’)2 form was inactive, indicating the relevance of the Fc gamma receptor to this ADCP (Fig. 7).
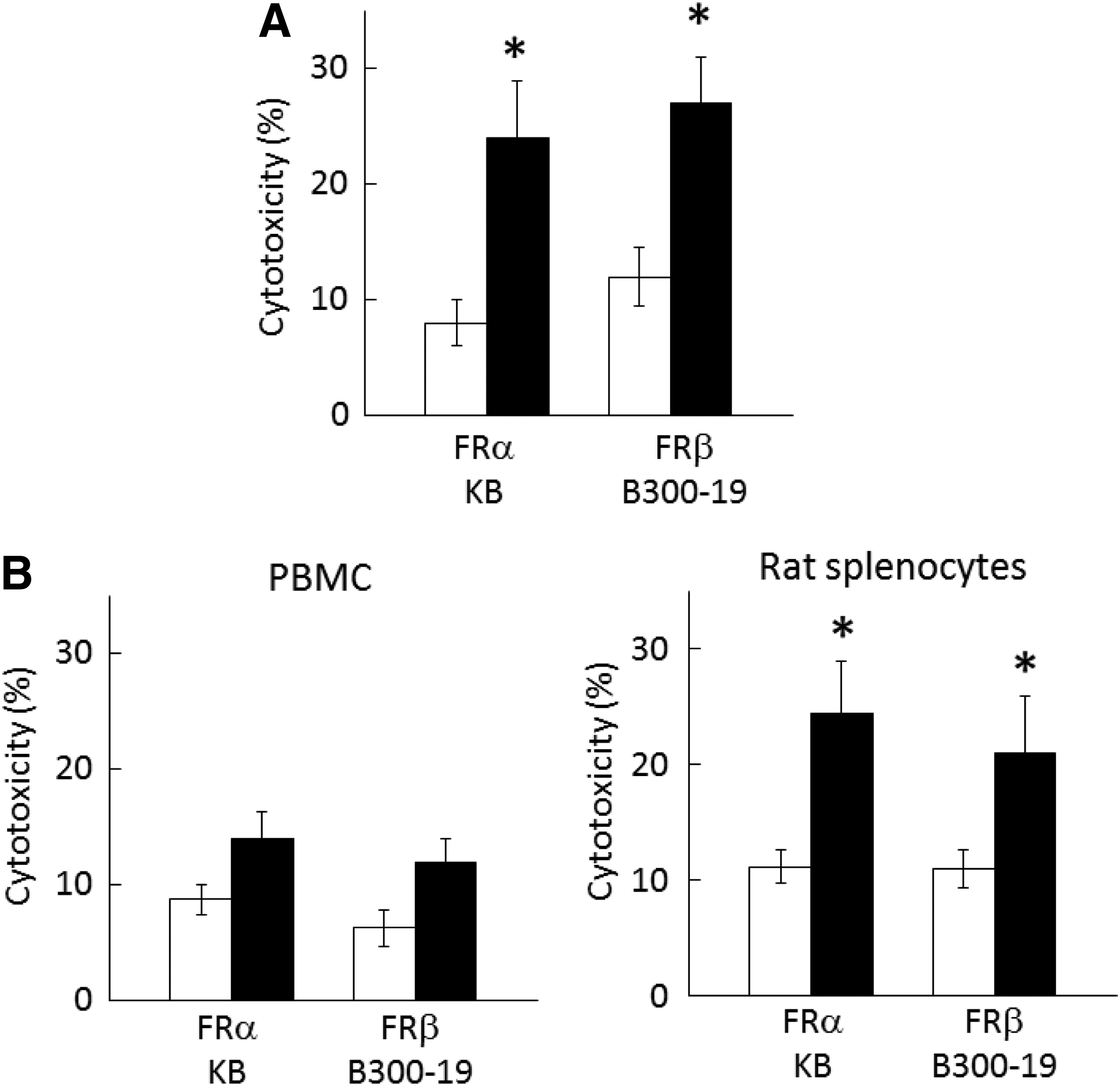
CDC and ADCC activity of clone 5 in FRα- and FRβ-expressing cells. (

ADCP activity of macrophages against FRα-expressing KB cells mediated by clone 5. (
Discussion
We obtained many anti-FRβ-producing hybridoma clones by immunizing human FRβ-transfected rat cell line into rats. Unexpectedly, we found that one clone reacts with FRα in addition to FRβ. Clone 5 (hereafter referred to as anti-FRαβ) reacted with FRα- and FRβ-expressing cell lines, a small fraction of peripheral blood monocytes, and most macrophages cultured with M-CSF. The immunohistochemical analysis showed that an anti-FRαβ stained cancer cells and a significant fraction of TAMs. The percentages of FRα positive cells in the serous type of ovarian cancer were lower compared to previous reports.(19,20) The small sample size in our study might have caused a slight bias. An anti-FRαβ also reacted with activated macrophages from the cynomolgus monkey and common marmoset. Further studies in nonhuman primates may provide useful information about the pharmacodynamics and toxicity of the anti-FRαβ MAb as a prelude to clinical trials.
An anti-FRαβ showed similar binding with high affinity to soluble FRα and FRβ. It has been reported that there is a significant extent of amino acid identity between FRα, FRβ, and FRγ and that FRγ may be present as the soluble form in serum but not on the plasma membrane of myeloid cells.(4) The binding of an anti-FRαβ to FRα- and FRβ-expressing cells was not inhibited in the presence of human serum. Although whether an anti-FRαβ reacts with FRγ has not been determined, the above finding supports that the MAb could target FRα- and FRβ-expressing cells in vivo, even in the presence of soluble FRγ.
Fc gamma-mediated effector functions such as CDC and ADCC have been suggested to be important for the in vivo activity of therapeutic antibodies.(21) An anti-FRαβ mediated CDC and ADCC in FRα- and FRβ-expressing cells, suggesting that this MAb could induce cytotoxicity in FRα-expressing cancer cells and FRβ-expressing TAMs in vivo. In addition, anti-FRαβ mediated ADCP of FRα-expressing KB cells by macrophages via the Fc gamma receptor. The cytotoxic or phagocytotic potential of a rat-derived anti-FRαβ in ADCC and ADCP using human cells may be increased by changing the rat Fc portion to a humanized form. Furthermore, a fully humanized anti-FRαβ will be required in clinical application for cancer.
It has been reported that TAMs can stimulate angiogenesis and enhance tumor cell invasion, motility, and intravasation. TAMs are also immunosuppressive, preventing tumor cell attack by natural killer and T cells during tumor progression.(22,23) There is strong evidence for specific subsets of TAMs as prognostic factors.(24–26) We reported that FRβ-expressing macrophages were prominent in the perivascular area of the tumor-invasive front and produced VEGF.(14,27) In fact, the increased number of FRβ-expressing TAMs was correlated with a poor prognosis in pancreatic cancer. Thus, an anti-FRαβ MAb that targets cancer cells and a subset of TAMs may become a more potent tool to inhibit cancer growth compared to either an anti-FRα or an anti-FRβ MAb. Folate-conjugated drugs delivered via FR-mediated cellular uptake have been shown to be promising anti-cancer therapeutics in preclinical animal models and early clinical trials.(28,29) Interestingly, a folate-conjugated mouse polyclonal immunoglobulin induced ADCC in FRα-expressing cells, inhibited tumor growth, and improved the survival rate in an animal model.(30) However, these folate-conjugated agents have not been well characterized with regard to their ability to target TAMs. Relative to anti-FR MAbs, FR-mediated cellular uptake of the folate-conjugated drugs may be inhibited by high folate serum concentrations. Furthermore, in addition to FR-expressing cells, folate-conjugated drugs may target RFC- and PFCT-expressing normal cells and cause detrimental side effects. One disadvantage using MAbs is their reduced penetration into tumors due to their high molecular weight compared to low molecular weight drugs. It is anticipated that administering a smaller anti-FRαβ MAb-derived polypeptide such as a single-chain or a single-domain MAb would result in an increase in tumor penetration.
In conclusion, this study suggests that the anti-FRαβ MAb described here is a promising therapeutic candidate for cancers in which macrophages promote tumor progression.
Footnotes
Author Disclosure Statement
The authors have no financial interests to disclose.