
Abstract
Respiratory syncytial virus (RSV) is the major cause of pulmonary inflammation in infants, young children, and immunocompromised adults. However, the RSV vaccine is not yet available commercially. The RSV-F glycoprotein mediates virus-host cell fusion, leading to syncytial formation; therefore, the RSV-F glycoprotein has been a treatment target for prevention and therapy of RSV infection. To produce the RSV-F-protein epitope-specific monoclonal antibody (MAb), BALB/c mice were immunized with a complex consisting of epitope peptide and MB-ODN 4531(O), encapsulated in a phosphatidyl-β-oleoyl-γ-palmitoyl ethanolamine (DOPE):cholesterol hemisuccinate (CHEMS) complex (Lipoplex(O)). Using conventional hybridoma technology, we obtained two clones able to produce antibodies reactive to two B-cell epitopes of RSV-F protein. Each anti-RSV-F glycoprotein MAb efficiently binds to each epitope. The F7-1A9D10 clone showed specific binding with RSV-F protein. There was no specific protein detected by Western blot analysis using F9 epitope-specific anti-RSV-F glycoprotein MAb (clone F9-1A6C8). However, based on confocal-image analysis, the antibody from the F9-1A6C8 clone showed specific binding with RSV-F protein. It is important that further study on possible applications for passive immunotherapy against RSV infection, such as therapeutic antibody production, is carried out.
Introduction
H
The RSV-F glycoprotein, which is the most effective of RSV proteins in provoking protective immunity, mediates virus-host cell fusion and leads to syncytial formation.(15) Therefore, the RSV-F glycoprotein has become the focus of efforts toward the prevention and therapy of RSV infection. A humanized, murine anti-RSV-F-glycoprotein monoclonal antibody (MAb), named palivizumab (Synagis), was developed to neutralize RSV.(16,17) This MAb was licensed for prophylaxis against RSV infection in high-risk infants and young children, and is commercially available in clinics. However, the application of palivizumab was not effective in cases of active or acute RSV infection.(17) To overcome the limitations of the palivizumab treatment, RSV-G glycoprotein has been investigated as a treatment target. Investigators have reported that anti-RSV-G-glycoprotein MAbs showed neutralization effects against RSV infection(18,19); however, this MAb is not yet licensed for clinical application against RSV infection.
MB-ODN 4531(O) was isolated as a natural CpG-DNA from the genomic DNA sequences of Mycobacterium bovis. MB-ODN 4531(O) with immunostimulatory activity contained three CpG motifs, which has been extensively studied by our research group as a vaccine adjuvant.(20) The adjuvant activity of MB-ODN 4531(O) was greatly enhanced by encapsulation with a complex containing phosphatidyl-β-oleoyl-γ-palmitoyl ethanolamine (DOPE) and cholesterol hemisuccinate (CHEMS); we named the whole complex Lipoplex(O).(21) Lipoplex(O) is an efficacious adjuvant for B-cell epitope screening using synthetic peptide and antibody production without carriers.(21) Previously, we screened and identified the B-cell epitopes (F7 and F9) of RSV-F glycoprotein by immunization with a complex consisting of each epitope peptide and Lipoplex(O).(22)
In this study, we produced an epitope-specific MAb recognizing RSV-F glycoprotein by immunizing mice with a complex of synthetic B-cell epitope of RSV-F glycoprotein (denoted F7 or F9 epitope) and Lipoplex(O). We then confirmed that the MAbs specifically recognized the RSV-F glycoprotein of RSV, as well as RSV-infected cells.
Materials and Methods
ODNs and reagents
The natural phosphodiester bond CpG-DNA, MB-ODN 4531(O) (synthesized by ST Pharm., Seoul, Korea), consists of 20 bases containing three CpG dinucleotides (underlined) flanked by specific base sequences: AGCAG
Cell culture and virus
HEp-2 cells were obtained from the American Type Culture Collection (ATCC, Manassas, VA) and Vero cells were kindly provided by Prof. Manseong Park (Korea University). The cells were maintained in Dulbecco's modified Eagle's medium (DMEM, Life Technology, Grand Island, NY) containing 10% fetal bovine serum (FBS, Life Technology, Grand Island, NY), 10 mM HEPES, 100 U/mL penicillin, and 100 mg/mL streptomycin. RSV A2 strain was kindly provided by Prof. Jun Chang (Ewha Womans University), and the virus was amplified on HEp-2 cells by a protocol reported previously.(23)
The recombinant H5N1 virus (PR8/H5Lo), the reassortant virus from segments of the A/Vietnam/1203/2004 (H5N1), and A/Puerto Rico/8/34 (PR8) (H1N1) influenza viruses were kindly provided by Prof. Manseong Park (Korea University).
Synthesis of B-cell epitope peptides
The B-cell epitope peptide of RSV A2 strain F protein (F7, 346AGSVSFFPQAETCKV360; F9, 393CKIMTSKTDVSSSVI407) was selected on the basis of its hydrophilicity value from Kyte and Doolittle,(24) its surface probability values from Emini and colleagues,(25) the α- and β-amphipathic values from Eisenberg and colleagues,(26) and the antigenicity index from Jameson and Wolf,(27) as described previously.(22) We also synthesized two typical variant sequences in several RSV strains, corresponding to the F7 (F7-1:346AGSVSFFPLAETCKV360, F7-2:346AGSVSFFPQADTCKV360) and F9 (F9-1:393CKIMTSK-ADVSSSVI407, F9-2:393CKIMTSKTDISSSVI407) epitope sequences.(22) The peptides were synthesized by Peptron (Daejeon, Korea) using the automated peptide synthesizer Peptron III-R24.
Preparation of epitope peptide and Lipoplex(O) complex
The complex of synthetic B-cell epitope peptide of RSV-F protein (F7 or F9) and Lipoplex(O) was prepared as described previously.(21) Briefly, DOPE and CHEMS were mixed well in ethanol (1:1 molar ratio) at room temperature. Nitrogen gas was injected into the mixture to make an ethanol-free lipid film. The DOPE and CHEMS mixture was re-suspended in a mixture of water-soluble MB-ODN 4531(O) and epitope peptide of RSV (F7 or F9 at 1:1 v/v), and then stirred vigorously at room temperature for 30 min. The pH of the complex of epitope peptide and Lipoplex(O) was adjusted to 7.0, and the complex solution was sonicated for 30 s. The complex solution was then filtered with a 0.22 μm filter and freeze-thawed three times with liquid nitrogen.
Mice and immunization of epitope peptide and Lipoplex(O) complex
Four-week-old female BALB/c (H-2b) mice were obtained from Orientbio (Seongnam, Korea) and maintained under specific-pathogen-free conditions in a controlled environment (20–25°C, 32–37% humidity). Animal studies were approved by the Institutional Animal Care and Use Committee of Hallym University (permit number Hallym 2010-10, Hallym 2013-55). The mice were injected intraperitoneally (i.p.) with the complex of each epitope (50 μg, F7 or F9) and Lipoplex(O), three times at 10-day intervals.
Production of mouse anti-RSV F protein monoclonal antibody
The spleens from immunized mice were removed, chopped by slide glass rubbing, and used for fusion to generate hybridomas.(28) The splenocytes were mixed with mouse SP2/0 myeloma cells at a 3:1 ratio in polyethylene glycol solution (PEG, Sigma-Aldrich) for fusion. After fusion, the cells were transferred to a 96-well plate (Nalgene Nunc International, Rochester, NY), and incubated at 37°C in 200 μL of 1% HAT (Sigma-Aldrich) in DMEM media containing 10% fetal bovine serum (FBS, Life Technologies, Grand Island, NY), 10 mM HEPES, 100 U/mL penicillin, and 100 μg/mL streptomycin. After colony formation, the culture media were collected and ELISA was performed to validate peptide-epitope-specific IgG production. Hybridoma cells were selected by two rounds of sub-cloning using a standard limiting dilution protocol.
Preparation of ascites and purification of anti-RSV F protein monoclonal antibody
To produce ascites, BALB/c mice were injected with 0.5 mL of pristane (Sigma-Aldrich) before 1–2 week injection with 1×106 hybridoma cells. After the ascites were filled in the peritoneal cavity, the mice were sacrificed under Zoletil 50+ Rompun anesthesia, and the ascites harvested with a 10 mL syringe and centrifuged at 3000 RPM and 4°C. The supernatants were harvested and stored in a deep-freezer at −80°C. The anti-RSV-F protein (F7 or F9-epitope)-specific MAb was purified from the ascites using protein-A agarose column chromatography and an automatic Econo system (Bio-Rad, Hercules, CA). The ascites were diluted with IgG binding buffer (Thermo Fisher Scientific, Waltham, MA), loaded on the column, and washed with 10 bead volumes of IgG binding buffer. After washing, the antibodies were eluted using IgG elution buffer (Thermo Fisher Scientific). The IgG fractions were collected and dialyzed with PBS buffer using a 20,000 MWCO Slide-A-Lyzer Dialysis Cassette (Thermo Fisher Scientific) four times at 6 h intervals.
Surface plasmon resonance analysis
The binding affinity of the anti-RSV F protein (F7 or F9-epitope)-specific MAb to the F7 or F9 epitope peptide was measured using a Biacore 3000 at 25°C. Biotinylated epitope peptides were captured on the flow cell surface of an SA sensor chip coated with streptavidin. A buffer solution containing biotin served as a negative control. The SPR data were evaluated with Biacore Bia evaluation software (v. 3.0, Biacore, GE Healthcare, Uppsala, Sweden). The sensor-gram data were also evaluated with the software. To show the binding affinity, graphs were reproduced using Microsoft Excel.
Antigen-specific Ig ELISA
Mouse sera were obtained from the eyeball veins before each injection, as well as after final injection, to detect antibody production after intraperitoneal injection with the complex of each epitope (50 μg, F7 or F9) and Lipoplex(O). Supernatants from the hybriboma cell culture were obtained to screen the peptide-specific IgG production. Ascites were also harvested from the peritoneal cavity. To measure the antibody titers of total IgG, 96-well immunoplates (Nalgen Nunc International, Penfield, NY) were coated with each of the RSV-F-protein epitopes (5 μg/well, F7 or F9) in 0.1% carbonate buffer. The plate was blocked with PBST containing 1% BSA for 1 h. The hybridoma culture supernatants, ascites, or purified MAb were added to the top row of the plate, and serial 1:4 dilutions in PBST were then placed into subsequent rows. The plates were incubated for 2 h at room temperature, washed with PBST three times, and then incubated with HRP-conjugated anti-mouse IgG antibody (Jackson Laboratory, Sacramento, CA) for 1 h. A colorimetric assay was carried out with a tetramethylbenzidine (TMB) peroxidase substrate solution (Kirkegaard and Perry Laboratories [KPL], Gaithersburg, MD), and stopped with TMB-stop solution (KPL). Absorbance was measured at 450 nm with a Spectra Max 250 microplate reader (Molecular Devices, Sunnyvale, CA).
To identify the isotype of MAb, 96-well plates were coated with each epitope peptide (F7 or F9), reacted with each MAb, and then incubated with HRP-conjugated anti-mouse IgG (each isotype) antibody (Jackson Laboratory). The reactivity was identified as described above.
To investigate whether the epitope-specific MAbs could bind with each corresponding epitope from other RSV strains, 96-well plates were coated with each epitope peptide: F7-1: 346AGSVSFFPLAETCKV360; F7-2: 346AG-SVSFFPQADTCKV360; F9-1: 393CKIM-TSKADVSSSVI407; or F9-2: 393CKIMTSKTDISSSVI407. Each MAb was added to the plates, and then incubated with HRP-conjugated anti-mouse IgG antibody (Jackson Laboratory). The reactivity was identified as described above.
Western blot analysis
To identify whether F-protein-epitope-specific MAb can detect RSV-F protein, viral particles of the RSV A2 strain were lysed with sample buffer composed of 10 mM HEPES (pH 7.4), 150 mM NaCl, 5 mM EDTA, 100 mM NaF, 2 mM Na3VO4, 10% NP-40 (Sigma-Aldrich), and one tablet of protease inhibitor (Roche, Indianapolis, IN); then separated by SDS-PAGE. The rH5N1 virus (PR8/H5Lo) was used as a negative control. The separated proteins were transferred to a pure nitrocellulose membrane (Pall Corp., Washington, NY) and blocked with 10 mL PBST containing 3% BSA for 0.5 h. Then 10 μg of each epitope-specific MAb was added and incubated for 2 h at room temperature. The membrane was washed with 10 mL PBST three times and incubated with PBST containing 5% skim milk (BD Biosciences, San Jose, CA). Finally, the membrane was incubated with HRP-conjugated anti-mouse IgG antibody (Jackson Laboratory, ratio 1:5000). The immunoreactive proteins were detected by the addition of an enhanced chemiluminescence solution (Amersham, Piscataway, NJ).
Immunofluorescence staining and confocal microscopy
Vero cells were cultured on 18 mm microscope glass cover slips in 12-well plates at a density of 2×104 cells/well. The cells were infected with RSV A2 strain with an MOI of 0.1 and incubated until cytopathic effect was observed. The infected cells were fixed with 4% paraformaldehyde and blocked with PBS containing 3% BSA and 0.1% Triton-X 100. The cells were incubated with each epitope-specific MAb against RSV-F protein or goat anti-hRSV antibody (Genway Biotech, San Diego, CA) and then incubated with Alexa Fluor 488-conjugated rabbit anti-mouse IgG or donkey anti-goat IgG (Life Technologies). The cells were also incubated with Hoechst no. 33258 (Life Technologies) to stain their nuclei. The cells were mounted with Fluoromount-G (Southern-Biotech, Birmingham, AL) and scanned with a Carl Zeiss LSM 710 (Carl Zeiss, Jena, Germany).
Results
Generation and purification of epitope-specific anti-RSV F glycoprotein monoclonal antibody
Previously, we identified two efficient B-cell epitopes from nine candidates of RSV-F glycoprotein. We also showed that active immunization with the peptide vaccine comprising a complex of RSV-F protein epitope (F7 or F9) and Lipoplex(O) improved the prophylactic effect against RSV infection.(22) These results provided evidence that epitope-specific anti-RSV-F glycoprotein antibody could be a potential therapeutic to prevent RSV infection. To extend this research, we produced epitope-specific anti-RSV-F glycoprotein MAb by immunization with a complex of RSV-F protein epitope (F7 or F9) and Lipoplex(O). BALB/c mice were injected three times at 10-day intervals with the complex of each RSV-F protein epitope (F7 or F9) and Lipoplex(O). After immunization of the mice, their sera were collected and production of each epitope-specific IgG was verified using ELISA. Mice with the highest titer of IgG recognizing each of the RSV-F-protein epitopes were selected. Mouse splenocytes were prepared and fused with mouse SP2/0 myeloma cells for generation of hybridoma cells. The hybridoma cells were screened, and two hybridoma cells producing RSV-F protein epitope-specific MAbs (F7-1A9D10 for F7 epitope, F9-1A6C8 for F9 epitope) were obtained (data not shown). Ascites containing epitope-specific MAbs were collected from the peritoneal cavity of BALB/c mice injected with the hybridoma cells. The RSV-F-protein epitope-specific MAbs were purified from the ascites using protein A agarose column chromatography. The purified MAbs were identified by SDS-PAGE and Coomassie Blue staining (Fig. 1A,D). The ELISA results showed that each epitope-specific MAb could bind with each epitope peptide (Fig. 1B,E). The purified MAbs were identified as IgG2b by ELISA using an IgG isotyping kit (Fig. 1C,F).

Production of RSV-F glycoprotein-specific MAbs. Hybridoma cells producing F7 epitope-specific (
Binding RSV-F protein epitope-specific monoclonal antibodies to synthetic epitope peptides
To measure the binding affinity of the anti-RSV-F protein F7 or F9 epitope-specific MAb, we performed SPR analysis using a Biacore 3000. We selected 1:1 binding with mass transfer as a fitting model for the ligand-analyte interaction. The F7 epitope-specific anti-RSV-F protein MAb reacted with the F7 epitope peptide, with a dissociation constant (Kd) of ∼2 nM and the F9 epitope-specific anti-RSV-F-protein MAb reacted with F9 epitope peptide with Kd of ∼ 0.02 nM (Fig. 2A,B).
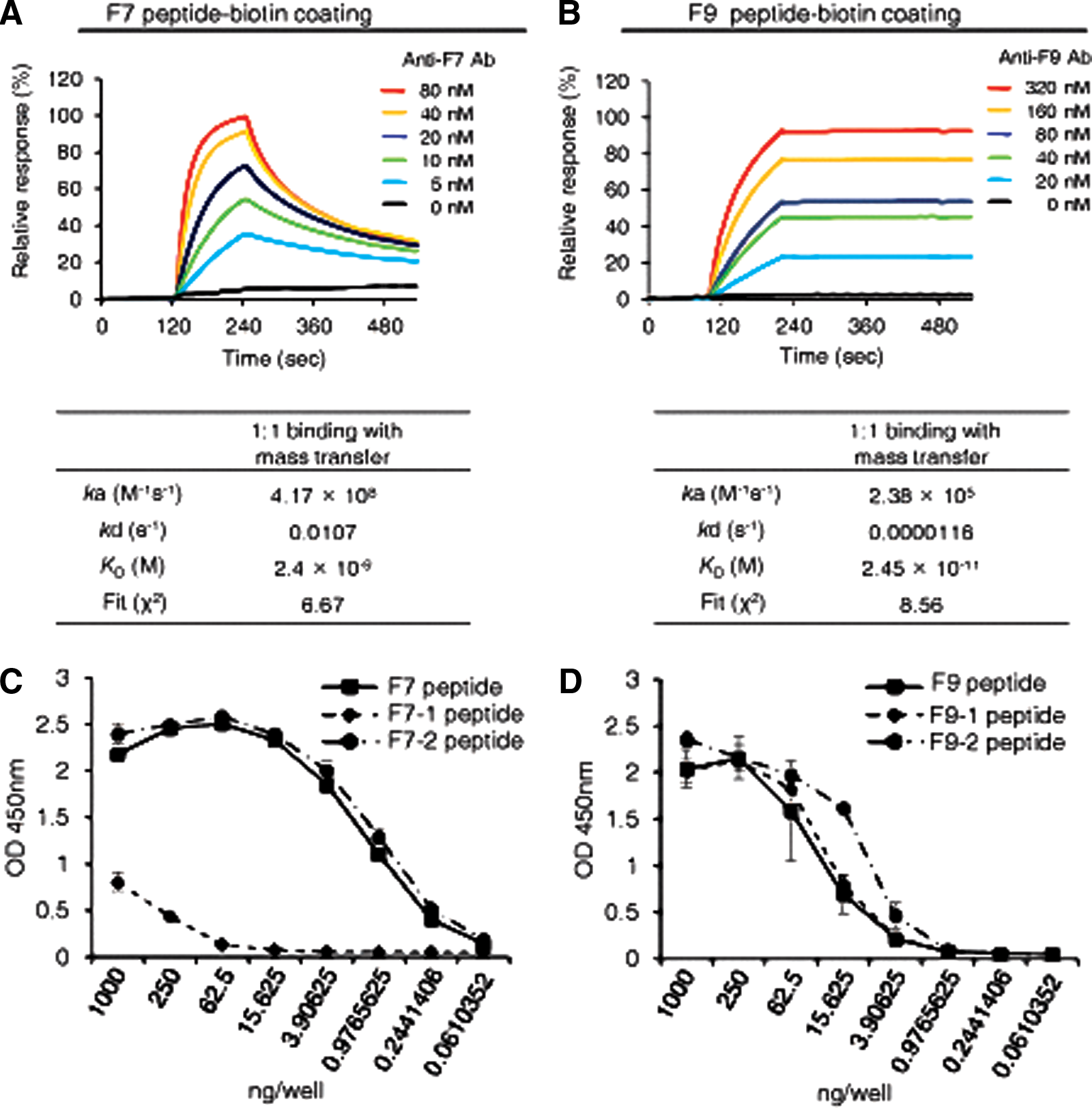
Binding of MAbs. (
Previously, we showed that the F7 and F9 epitopes are highly conserved among the RSV strains reported until this point, with only one amino acid difference.(22) Therefore, using ELISA, we next determined whether the epitope-specific MAbs could bind with each corresponding epitope. The F7 epitope-specific anti-RSV-F glycoprotein MAb (clone F7-1A9D10) efficiently bound to F7 and F7-2 epitopes, but the MAb could not bind to the F7-1 epitope (Fig. 2C). The F9 epitope-specific anti-RSV-F glycoprotein MAb (clone F9-1A6C8) showed specific binding activity to every F9 epitope (Fig. 2D).
Western blot analysis and immunoprecipitation analysis with RSV-F protein epitope-specific monoclonal antibodies
To investigate whether the purified RSV-F-protein-epitope-specific MAbs could recognize RSV-F glycoprotein in the RSV A2 strain, Western blot analysis was performed with each clone of RSV-F-protein epitope-specific MAbs. The viral proteins of the RSV A2 strain were analyzed using SDS-PAGE and subjected to Western blot analysis, along with the proteins of the recombinant H5N1 influenza virus (PR8/H5Lo) as a negative control. The F7-1A9D10 clone showed specific binding with RSV-F protein without recognition of recombinant H5N1 influenza virus proteins (Fig. 3). However, no specific band was found in the Western blot analysis with F9 epitope-specific anti-RSV-F glycoprotein MAb (clone F9-1A6C8) (data not shown).
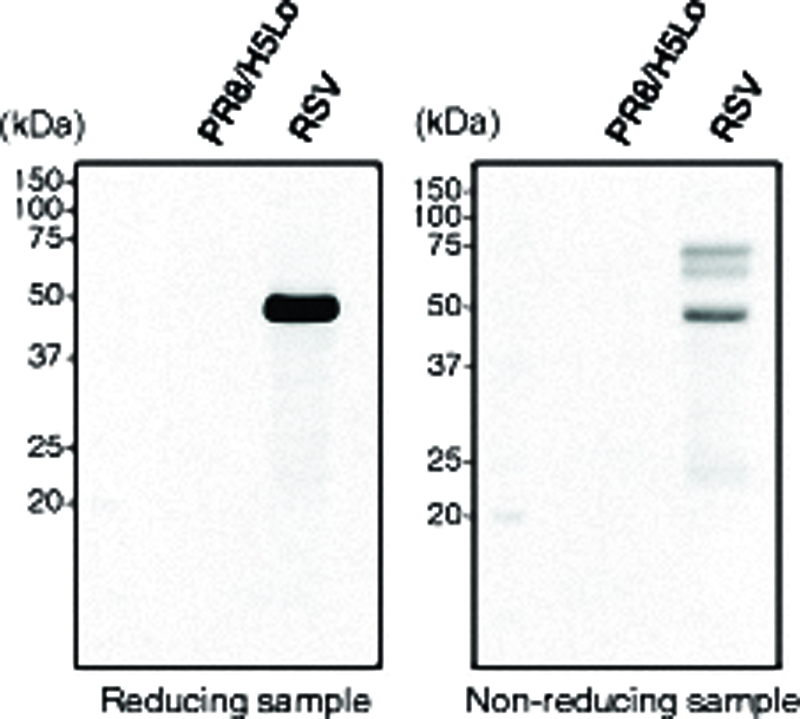
Western blot analysis. Viral particles of RSV A2 strain were prepared and separated by SDS-PAGE, and Western blot analysis was performed using F7 epitope-specific anti-RSV F glycoprotein MAb. Proteins of recombinant H5N1 influenza virus (PR8/H5Lo) were used as negative control.
Confocal image analysis of RSV-F-protein epitope-specific monoclonal antibody
To identify the recognition of RSV in Vero cells by each RSV-F-protein epitope-specific MAb, the cellular localization of RSV was examined by immunofluorescence staining and confocal imaging. The Vero cells were infected with the RSV A2 strain for 72 h, and the cells were then incubated with each epitope-specific MAb against RSV-F protein, or goat anti-hRSV antibody. As shown in Figure 4, the confocal images reveal that RSV-F-protein expression was detected by each RSV-F-protein epitope-specific MAb, with higher reactivity found in the case of F9 epitope-specific MAb.

Immunostaining and confocal image analysis. Vero cells were incubated with RSV A2 for 72 h, and detection of RSV-F protein by MAbs was analyzed by indirect immunofluorescence staining and confocal microscopy. RSV-F protein was stained with F7 epitope-specific, F9 epitope-specific anti-RSV-F glycoprotein MAb, or commercially available goat anti-RSV polyclonal antibody, and then detected with FITC-conjugated secondary antibody (green). Nuclei were stained with Hoechst No. 33258 (blue). Scale bars, 100 μm.
Discussion
CpG-DNA (i.e., synthetic oligodeoxynucleotides and bacterial chromosomal DNA composed of CpG dinucleotides flanked by specific nucleotide sequences) has immune-stimulatory function in the innate immune system.(29,30) Many investigators have shown adjuvant activity for CpG-DNA. For example, CpG-DNA contributes to the production of antibodies against conventional protein antigens such as ovalbumin.(31,32) Previously, we developed Lipoplex(O) to enhance the adjuvant activity of CpG-DNA. Lipoplex(O) has significant adjuvant activity for the production of antibodies against protein antigens such as ovalbumin, hen egg lysozyme, and SUMO-1.(33) We extended the application of Lipoplex(O) as an adjuvant activity to produce peptide epitope-specific antibody without carriers. We confirmed that immunization with a complex of epitope peptide and Lipoplex(O) significantly boosts the production of antibodies against epitope peptides from hepatitis B virus,(34) influenza virus,(35) RSV,(22) and tumor antigen.(36) Furthermore, we showed that immunization with a complex of epitope peptide and Lipoplex(O) protected mice from influenza virus infection,(35) RSV infection,(22) and hepatocellular carcinoma implantation.(36) Here, we produced two clones of anti-RSV-F glycoprotein MAbs by immunizing mice with a complex of synthetic B-cell epitope of RSV-F glycoprotein: clone F7-1A9D10 against F7 epitope and clone F9-1A6C8 against F9 epitope (Fig. 1).
RSV infects epithelial cells of the respiratory tract, which is the major site of its replication, and is enveloped by a lipid bilayer containing F and G glycoproteins. The RSV-F glycoprotein is synthesized in the host cytoplasm as an inactive precursor (F0) with a molecular weight of ∼ 60 kDa.(15,37) The F glycoprotein precursor is cleaved by furin-like proteases to produce two disulfide-linked subunits: F2 from the N-terminus (14–16 kDa) and F1 from the C-terminus (44–46 kDa).(38) The processed RSV-F glycoprotein is matured for formation of homotrimers.(39) The mature RSV-F glycoprotein triggers membrane fusion between RSV and the host cell membrane. The F glycoprotein also mediates cell-cell fusion, inducing formation of syncytia. Therefore, production of anti-RSV-F antibody to neutralize the RSV is important for protection and therapy against RSV infection.
Here, we produced two clones of monoclonal antibodies against RSV-F glycoprotein. The clone F7-1A9D10 was reactive with F7 epitope, based on SPR analysis and Western blot analysis (Figs. 2 and 3). However, the signal in the confocal microscopy after immunostaining was weaker than that for the other clone F9-1A6C8. On the other hand, clone F9-1A6C8 was highly reactive with F9 epitope, with very low Kd (∼ 0.02 nM), and showed strong immunoreactivity in the confocal images. However, it cannot be used for Western blot analysis. Therefore, we have to select clones suitable for specific purposes. While clone F9-1A6C8 was highly reactive with other F9 peptides from different viral strains, clone F7-1A9D10 showed biased affinity (Fig. 2). This suggests, therefore, that F9-1A6C8 would be a better choice for diagnostic and therapeutic purposes.
In conclusion, we selected two specific B-cell epitopes of RSV-F glycoprotein, and produced two clones of monoclonal antibodies specific to RSV-F glycoprotein. Further studies are needed to investigate the possible application of these antibodies as therapeutics against RSV infection through passive immunization.
Footnotes
Acknowledgments
This research was supported by the National Research Foundation of Korea funded by the Ministry of Science, ICT & Future Planning in the Republic of Korea (2013R1A2A2A03067981, 2013M3A9A9050126, 2014M3C1A3051473, 2014047905).
Author Disclosure Statement
The authors have no financial interests to disclose.