
Abstract
In human cancers, over-expression of HLA-E is marked by gene expression. However, immunolocalization of HLA-E on tumor cells is impeded by the HLA-Ia reactivity of commercial anti-HLA-E monoclonal antibodies (MAbs). So there was a clear need to develop monospecific anti-HLA-E MAbs for reliable immunodiagnosis of HLA-E, particularly considering the prognostic relevance of HLA-E in human cancer. HLA-E overexpression is correlated with disease progression and poor survival of patients, both of which are attributed to the suppression of anti-tumor activity of cytotoxic T cells mediated by HLA-E. The suppression mechanism involves the binding of HLA-E-specific amino acids located on the α1 and α2 helices of HLA-E to the inhibitory receptors (CD94/NKG2a) on CD8+ T lymphocytes. An anti-HLA-E MAb that recognizes these HLA-E-specific sequences can not only be a monospecific MAb with potential for specific immunolocalization of HLA-E but can also block the sequences from interacting with the CD94/NKG2a receptors. We therefore developed several clones that secrete such HLA-E-specific MAbs; then we assessed the ability of the MAbs to bind to the amino acid sequences interacting with the CD94/NKG2a receptors by inhibiting them from binding to HLA-E with peptides that inhibit receptor binding. Elucidation of the immunomodulatory capabilities of these monospecific MAbs showed that they can induce proliferation of CD8+ T cells with or without co-stimulation. These novel MAbs can serve a dual role in combating cancer by blocking interaction of HLA-E with CD94/NKG2a and by promoting proliferation of both non-activated and activated CD8+ cytotoxic αβ T cells.
Introduction
M
Binding of HLA-E to the inhibitory receptors CD94 and NKG2A on both CD8+ CTLs and NKT cells may impede the cell-mediated anti-tumor activity. The interaction between HLA-E and the inhibitory receptors involves the binding of amino acids located on the α1 and α2 helices of HLA-E to specific amino acids on CD94 and NKG2A, the α1 and α2 of HLA-E being, respectively, (R65D69Q72I73R75V76R79) and (E167D163A159H156E155S152); CD94 and NKG2A being, respectively, (Q112Q113F114L162D163E164N170) and (R137P171S172K199Q212K217).(24) This interaction is attributed to loss of the anti-tumor activity of CD8+ CTLs as well as that of NK or NKT cells. The amino acid sequences on HLA-E recognized by the inhibitory receptors are unique and specific for HLA-E. Synthetic peptides of these amino acid sequences can be used to ascertain the specific binding affinity of the anti-HLA-E MAbs.(25)
The identification and localization of HLA-E on the surface of cancer cells 1 and serum-soluble HLA-E(26) is based on immunostaining with any one of these commercially available anti-HLA-E MAbs: MEM-E/02, -E/06, -E/07, -E/08, or 3D12—on the assumption that they are specific for HLA-E. Paradoxically, all these MAbs react not only with HLA-E but also with HLA-Ia and/or HLA-Ib alleles.(27,28) Since the reactivity of these HLA-E MAbs was inhibited by the peptides (117AYDGKDYLT125, 126LNEDLRSWTAV136, and 137DTAAQI142) shared by both HLA-Ia and HLA-Ib alleles,(27,28) these anti-HLA-E MAbs were confirmed as being polyspecific and not suitable for reliable localization of HLA-E on tumor cell surfaces; there was a need both to generate anti-HLA-E MAbs that can bind only to amino acid sequences (epitopes) unique or specific for HLA-E and to validate the expression of HLA-E on the cell surface of cancer cells or virally infected cells. In this study, the binding of anti-HLA-E MAbs to HLA-E-specific peptides present on α1 and α2 helices was determined by using synthetic peptides for dosimetric inhibition of the MAbs' binding to HLA-E after ascertaining that the binding was restricted to HLA-E.
Both the poor prognosis of cancer patients with overexpressed HLA-E(21–23,29,30) and the ability of monospecific anti-HLA-E MAbs to bind at the site of the epitopes of CD94 and NKG2A on HLA-E suggest the possible use of monospecific anti-HLA-E MAbs to mask the binding sites of the inhibitory receptors on HLA-E. Such blocking of HLA-E by monospecific MAbs may help restore the anti-tumor efficacy of CD8+ T cells that was lost from the interaction of inhibitory receptors and HLA-E. It is well known that restoration and clonal expansion may enable CD8+ T cells to release molecules that activate lytic pathways on virally infected tumor and allograft (organ transplant) cells.(42)
While transplant clinicians aim to suppress CD8+ T cells to prevent allograft rejection, oncologists and virologists strategize to activate and proliferate CD8+ cells to kill target cells. Anti-CD3 MAb is used to expand CTLs in vitro, but it is not specific since it allows proliferation of all other categories of CD3+ T cell populations (CD4+/CD8-, CD4+/CD8+ and CD4-/CD8-).(4) A strategy to induce specific proliferation of CTLs without activating other T cell subsets in vivo is needed when treating cancer and infection. To that end, we noted earlier that the polyspecific anti-HLA-E MAbs, TFL-006 and TFL-007—but not the monospecific anti-HLA-E MAb, TFL-033—suppressed activated CD4+ as well as CD8+ CTLs much better than did intravenous immunoglobulin.(41,42) While the polyspecific anti-HLA-E MAbs recognized the peptide sequences 117AYDGKDYLT125, 126LNEDLRSWTAV136, and 137DTAAQI142 shared by HLA-A, -B, -Cw, -F, and -G),(27,28,43–47) the monospecific MAbs recognized HLA-E-specific amino acid sequences on the α1 and α2 helices—respectively, 65RSARDT70 and 153AESADNSKQES143.(25) The differential peptide recognition by polyspecific and monospecific anti-HLA-E MAbs may contribute to the different functional capabilities of anti-HLA-E MAbs.
We hypothesized that the monospecific anti-HLA-E MAbs, although not capable of suppressing the CD3+, CD4+, or CD8+ T cells, may promote proliferation of T cells. If such proliferation of CD8+ T cells is consistently mediated by a number of monospecific anti-HLA-E MAbs, not only would the hypothesis be validated but these monospecific anti-HLA-E MAbs may have significant immunotherapeutic relevance. The hypothesis was tested by sequential investigations:
1. Several clones secreting anti-HLA-E MAbs were generated after immunization with two different alleles of recombinant HLA-E heavy chains (EG107 and ER107) in a mouse model; 2. MAb monospecificity was determined by examining their dose-dependent binding to microbeads coated with different alleles of HLA class Ia (HLA-A, -B, and -Cw) and Ib (HLA-E, -F, and -G); 3. Binding specificity of a selected clone was determined using the HLA-E-specific or unique peptide sequences that are also the target site of inhibitory receptors of CTLs; 4. Immunodiagnostic potential was assessed by comparing the immunostaining of two different human cancer tissue microarrays with the monospecific MAbs and a commercial, commonly used polyspecific anti-HLA-E MAb (MEM-E/02); 5. The influence of the monospecific anti-HLA-E MAbs on non-activated and PHA-activated CD8+ T cells was evaluated.
The results showed the capability of the monospecific anti-HLA-E MAbs of promoting proliferation of CD8+ T cells in addition to facilitating specific immunodiagnosis of HLA-E on tumor cells.
Material and Methods
Production of MAbs
The immunization protocols, development of hybridoma, selection of clones, isotype characterization, and preparation of culture supernatants and ascites used to produce HLA-E monospecific and polyspecific MAbs have been described elsewhere.(25,43–46) The recombinant (r) HLA-EG107 and HLA-ER107 (10 mg/mL in MES buffer) heavy chains, devoid of β2-microglobulin (β2m), obtained from the Immune Monitoring Lab, Fred Hutchinson Cancer Research Center (University of Washington, Seattle, WA) were used to immunize the mice (Table 1A).
The experiment used human-lg-absorbed Goat anti-mouse F(ab′)2 region Abs specific to (H+L) of Isotypes (Southern Biotech). Concentration of all secondary antibodies was 0.5 mg/ml; 50 μl was used after diluting 1/100.
Immunoassay to characterize the specificity of MAbs
To determine the mono- or polyspecificity of the MAbs, multiplex Luminex®-based immunoassay (One Lambda, Canoga Park, CA) was used, as previously described.(27,28,43–46) The purified culture supernatants and ascites of the MAbs were serially diluted with PBS (pH 7.2) before mixing with microbeads coated with HLA-I alleles. Using dual-laser flow cytometry (Luminex xMAP® multiplex technology, Luminex Corp., Austin, TX), single antigen assays were carried out for data acquisition and analysis, as reported earlier.(43–49) The single recombinant HLA-Ia antigens in LS1A04-lot 007 were used for screening the MAbs. This lot contains 31 HLA-A, 50 HLA-B, and 16 HLA-Cw allelic molecules. In addition to recombinant HLA-E, the heavy chains of HLA-F and HLA-G (also from the Immune Monitoring Lab) were individually attached by a process of simple chemical coupling to 5.6 μm polystyrene microspheres with infrared fluorophores. The HLA-Ia microbeads have built-in control beads: human IgG–coated positive beads and serum albumin–coated negative beads. For HLA-Ib, both positive and negative beads were added separately. Antigen density on the single antigen beads coated with HLA-Ia molecules was determined using MAb W6/32, per the manufacturer's instructions (One Lambda). The binding level (antigen density) of HLA-E, -F, and -G to the beads was assessed by using HLA-Ib–reactive polyspecific TFL-006 and TFL-007 MAbs on the HLA-E, -F, and -G coated single antigen beads and checked to see if their mean fluorescent intensity (MFI) is comparable; the epitope targeted by TFL-006 and TFL-007 was found in the heavy chains of HLA-E, -F, and -G coated on the beads. The data were published elsewhere.(44,46) The MAbs were used at different dilutions (see Figs. 1 and 2). Secondary fluorescence-labeled anti-mouse polyclonal Abs were diluted 1/100 (anti-mouse IgG [H+L], cat. no. 115-116-146, protein concentration 0.5 mg/mL; Jackson ImmunoResearch Laboratories, West Grove, PA). For identifying the isotypes, anti-Fc secondary mouse anti-isotype Abs were used at concentration 0.5 mg/mL and dilution 1:100; they were: human-absorbed anti-mouse IgG1 (cat. no. 1070-095); IgG2a (cat. no. 1080-095); IgG2b (cat. no. 090-095); IgG3 (cat. no. 100-095; all from SouthernBiotech, Birmingham, AL). Data generated with Luminex Multiplex Flow cytometry (LABScan 100) were analyzed using the same computer software and protocols reported earlier.(43–49)
(

Titrimetric profiles of ascites compared with culture supernatant of MAb TFL-185 (
Peptide inhibition assays
In contrast to the peptides used for the MAbs, two different HLA-E-restricted peptides—65RSARDTA71 and 143SEQKSNDASE152—were used to inhibit the binding of monospecific MAbs, the source and characteristics of the peptides having been described earlier.(25) Various concentrations of peptides (serially diluted from the initial concentration of 100μg to 100 μL) were added to the MAbs (7 μL), further diluted by adding 14 μL PBS-BSA (pH 7.0; final dilution 1/1200), then exposed to 2 μL of beads. PE-conjugated anti-mouse IgG was used for the LabScreen assay, and each experiment was done in triplicate.
Tissue microarrays
Three tissue microarrays (TMA, US Biomax, Rockville, MD) were carefully selected for immunohistochemistry: they were TMA that included 70 primary melanoma tissues and 10 normal tissues plus information on pathological diagnosis, TNM stages, and cancer grade (Fig. 1A). The cores of all three TMA were 1.5 mm in diameter and 5 μm thick.
Immunohistochemistry
Details of the immunohistochemistry performed on the TMA are given elsewhere.(25)
Isolation and processing of T lymphocytes
On the day of the experiments, 60 mL of whole blood from a healthy normal male donor 2 was collected after obtaining the donor's informed consent and Institutional Committee approval. Human T lymphocytes (including both CD4+ and CD8+) were isolated from the whole blood using Ficoll™-Hypaque (GE Healthcare Bio-Sciences, Piscataway, NJ) to recover peripheral blood.(43,45) The isolated lymphocytes were separated into two lots, one treated with phytohaemagglutinin (PHA, cat. no. L-1800-100; EY Laboratories, San Mateo, CA) at a final concentration of 2.25 μL/mL, the other with no PHA, and both were cultured in 96-well tissue culture plates containing serum-free AIM V® Medium (cat. no. 12055-083; Life Technologies-Gibco, Grand Island, NY) with 1% HEPES, optimal pH range between 7.2 and 7.5. The MAbs were added to cells in culture within 2 h of PHA being added (the total volume was adjusted to 200 μL).(43,51) The number of cells after PHA activation or exposure to MAbs was measured by flow cytometry after staining the cells with PE-labeled anti-CD4 and PerCP-labeled anti-CD8 MAbs.
Monospecific anti-HLA-E MAbs upregulate non-activated and PHA-activated T cells
We used the culture supernatants from individual clones that were cultured—as described elsewhere(43–45)—in a medium containing RPMI 1640 w/L-glutamine and sodium bicarbonate (cat. no. R8758; Sigma-Aldrich, St. Louis, MO), 15% fetal calf serum, 0.29 mg/mL L-glutamine/Penn-Strep (cat. no. 400-110; Gemini-Bio, MedSupply Partners, Atlanta, GA), and 1 mM sodium pyruvate (cat. no. S8636; Sigma). Five hundred μL aliquots of the supernatants stored at −20°C were used in the investigation. Isotypes of all the MAbs were characterized, and no IgM Abs were detected. The IgG in the culture supernatant (400μL) was purified by passing each aliquot of supernatant through a Protein G Spin Kit (0.2 mL) (cat. no. 89949; Thermo Scientific, Waltham, MA). The TFL IgG MAbs were purified with in-house buffers (binding buffer, 50 mM sodium acetate, pH 7; elution buffer, 100 mM glycine, pH 2.8; and neutralizing buffer, 1M Tris-HCl, pH 8.5) without any NaN3. Commercial buffers were avoided since they contain 0.02% sodium azide.
The purified Abs were lyophilized overnight at 37°C using Speedvac (Thermo Scientific). The purified culture supernatants and ascites were tested for HLA reactivity. The protein concentrations of the MAbs were measured with a BioPhotometer (Eppendorf, Hauppauge, NY). In particular, as noted, the purified MAbs were diluted with AIM V Medium plus 1% HEPES, which had an optimal pH range between 7.2 and 7.5. Purified, sodium azide-free supernatants, ascites, and purified/lyophilized ascites were used to study the immunomodulation of T cells. All preparations used for immunomodulation studies were free of sodium azide.
Measurement of blastogenesis of activated T cells
Blastogenesis of CD4-/CD8+ T cells was determined by counting the lymphoblasts after culturing purified lymphocytes from donors for 72 h with PHA or without it. The CD8+ positive T cells include both CD3+ and CD3- lymphocytes, implying that it contains both CD8+ cytotoxic T cells and CD8+ NK (also known as NKT) cells. Lymphoblasts (group 3) were recognized by flow cytometry based on size (side scatter) and granularity (forward scatter).(43,45) In the tables and figures of flow cytometric profiles, three groups of T cells were identified for CD4+ and CD8+ T lymphocytes. They are groups 1 and 2, which comprise subsets of resting lymphocytes, and group 3, which comprises lymphoblasts.(43)
Statistical analysis
Primarily, pair-sample or equal-variant t-tests were carried out in this investigation using a graphic website (
Results
Characterization of anti-HLA-E MAbs
Diversity of isotopes of anti-HLA-E MAbs
In all, 258 hybridomas were generated from the spleens of mice immunized with the heavy chains of rHLA-EG107 and rHLA-ER107 (Table 1B). The HLA-EG107 generated 87 clones per single mouse whereas HLA-ER107 yielded 171 clones after immunizing two mice. While both HLA-E alleles are immunogenic, the immunogenicity of HLA-ER107 may be more immunogenic than HLA-EG107. The glycine at 107 of HLA-EG107 is commonly shared with HLA-F, HLA-G (Table 3), and several other HLA-Ia alleles, whereas arginine 107 is extremely rare among HLA-I, with two exceptions, HLA-A*01:01:01:02N and HLA-B*15:30. Such rarity probably accounts for the better immunogenicity of HLA-ER107 in both mice and humans.
The most common isotype of the IgG Abs was IgG1 (n=138); the other isotypes were IgG2a (n=48), IgG2b (n=53), IgG3 (n=8), and unknown (n=11). The reactivity of the anti-HLA-E MAbs differed markedly in their recognition of HLA-Ia and Ib alleles (Table 1C).
Diversity of HLA-Ia and -Ib reactivity of anti-HLA-E MAbs
Eight different groups of anti-HLA-MAbs can be broadly identified by their reactivity to HLA-Ia and Ib alleles as presented in Table 1C. They can be categorized as HLA-Ia-nonreactive (groups 1–4) and HLA-Ia-reactive (groups 5–8). Each group had HLA-E-specific Abs (groups 1 and 5) or Abs specific for HLA-E and HLA-F (groups 2 and 6), for HLA-G (groups 3 and 7), or for HLA-F and HLA-G (groups 4 and 8). Most importantly, group 1 consisted of HLA-monospecific MAbs and group 8 consisted of HLA-E polyreactive MAbs. The majority of commercial anti-HLA-E MAbs, which are claimed to be specific for HLA-E (e.g., MEM-E/02, MEM-E/06, MEM-E/07, MEM-E/08, and 3D12) fall into HLA-polyreactive groups 5–8. Using these polyreactive commercial MAbs for immunodiagnosis may not yield true expression of HLA-E on tissues unless they are used in conjunction with the monospecific HLA-E MAbs, as was carried out earlier (vide infra).(25)
MFI and isotypes of anti-HLA-E monospecific MAbs
Table 2 shows HLA-E monospecific MAbs ranked by mean fluorescent intensity (MFI), high to low. Six monospecific MAbs with MFI <1000 are not included in the table. The table shows that several MAbs demonstrated a higher MFI with isotype-specific MAbs.
anti-HLA-E monospecific monoclonal antibodies with overall MFI >1000 are not included
They rarely reacted with other classical or non-classical HLA. Of the total number of such Abs, only 24 had a MFI >1000. The HLA reactivity of any MAbs with a MFI <500 is considered negative; the MFI of any HLA-A, -B, or -Cw alleles rarely reached 400.
Titration of culture supernatant, ascites, and protein G-purified HLA-E monospecific MAbs
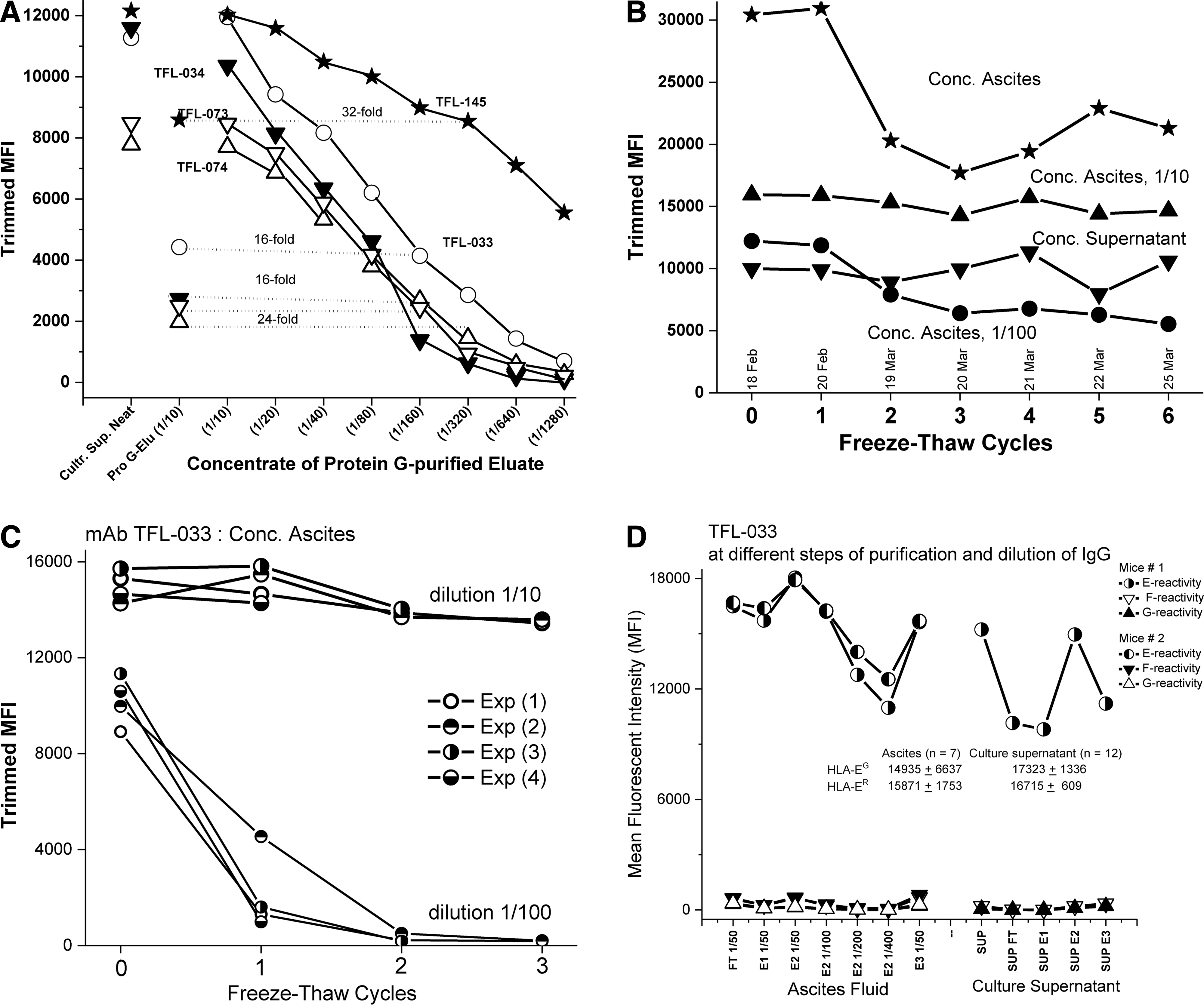
Figure 1A shows the reactivity of five different HLA-E monospecific MAbs (TFL-033, -034, -073, 073, and -145) assessed by the MFI of the culture supernatant of the clones, by the MFI of the IgG of the culture supernatant before and after eluting IgG from the protein G columns, and by the dosimetry of the concentrate of the protein G-eluted culture supernatant (neat and at different dilutions). Figure 1B shows the relative resistance of MAb TFL-033 to repeated freeze-thaw cycles, indicating the stability of TFL-033 in concentrated culture supernatant and ascites as well as at two different dilutions (1/10 and 1/100) of ascites after six freeze-thaw cycles. The high MFI of the concentrated ascites before the freeze-thaw cycles was due to aggregation caused by a high concentration of IgG Abs—an inference supported by the fact that dye-conjugated anti-IgG Ab (One Lambda) bound to heavy and light chains, not just to the Fc portion of IgG, so one could detect the aggregates of Abs bound to beads. Further proof is the decrease observed in MFI when the concentrated ascites were analyzed after centrifugation. The stability of MAb TFL-033 in ascites at 1/10 and 1/100 dilutions was validated by three freeze-thaw cycles (Fig. 1C). That stability tends to decline at higher dilutions. Figure 1D shows the monospecific reactivity of TFL-033 in ascites, in culture supernatant, and after protein G purification of IgG from culture supernatants; flow-through and eluates 1 to 3 at different dilutions consistently showed that none of the eluates or flow-through reacted to HLA alleles other than HLA-E. Figure 1C shows that the MAb TFL-033 reacted equally with both HLA-EG107 and HLA-ER107. Figure 2 compares the titrimetric profile of ascites and culture supernatant of MAbs TFL-185 and TFL-033.
Luminex validation of specificity of MAb TFL-033 versus commercial MAbs used for immunodiagnosis of HLA-E in cancer tissues
The LABScreen panel of color-coded beads were immunostained with different dilutions of TFL-033 and commercial HLA-E MAbs (MEM-E/02 and 3D12). MEM-E/02 and 3D12 stained both HLA-EG107 and HLA-ER107, but not HLA-F or HLA-G. They did, however, react dosimetrically with 25 HLA-A, 50 HLA-B, and 16 HLA-Cw beads (Fig. 3). This was in striking contrast to culture supernatants and ascites ofTFL-033, which reacted only with HLA-E alleles, confirming their monospecificity.
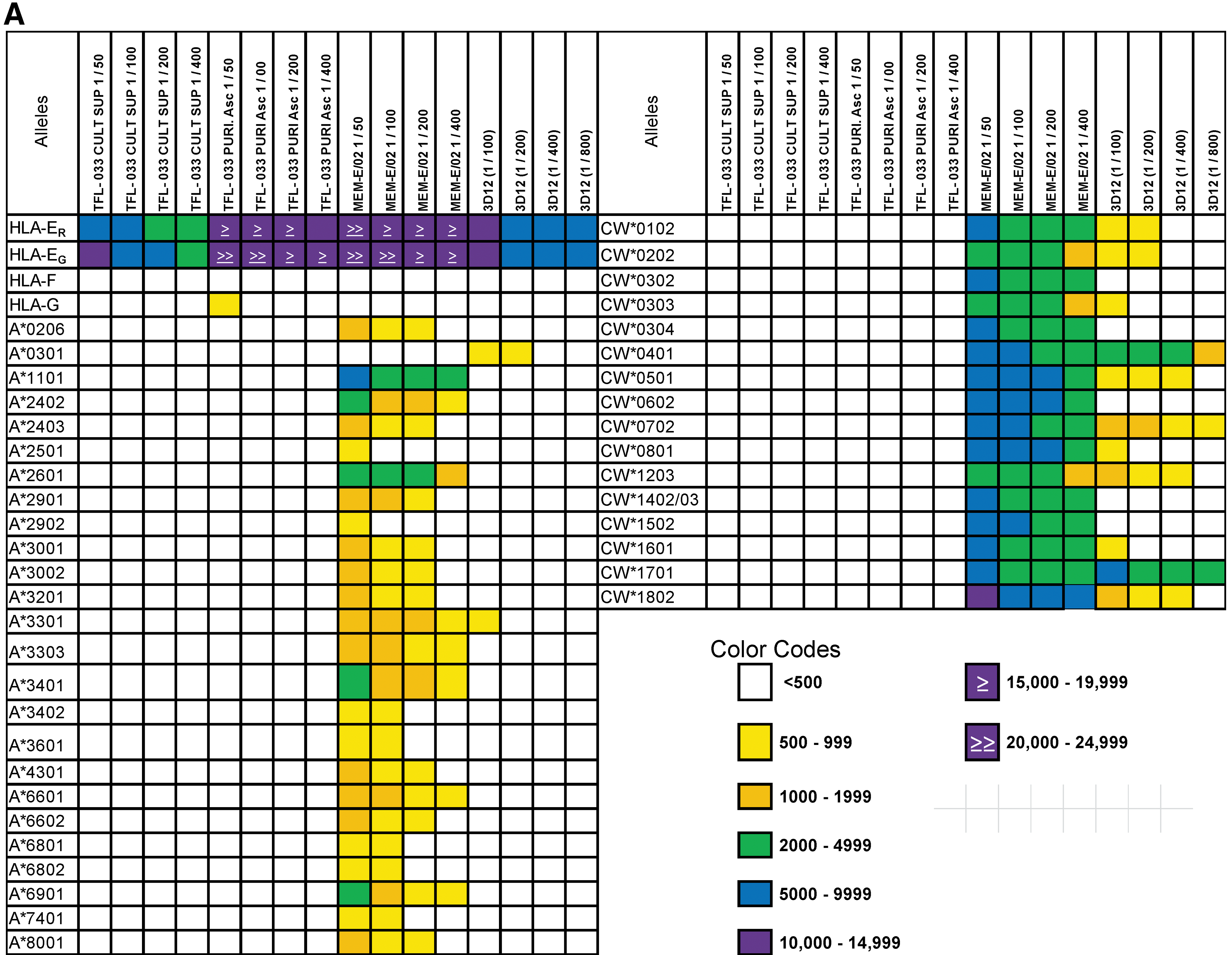
The much greater HLA reactivity of MAb TFL-033 at different dilutions of culture supernatant and ascites compared with that of MEM-E/02 and 3D12, which are extensively used to localize HLA-E in tumor tissues on the faulty assumption that they are specific for HLA-E. (
HLA-E-specific peptide sequences and peptide inhibition of HLA-E monospecific MAb
In comparing the amino acid sequences of HLA-E with the loci of HLA-A, -B, -Cw, -F, and -G, it was noted that at least five peptide sequences (the amino acid sequences shown in Table 3) were restricted to HLA-E. Two sequences were located on α1 helices, two on α2 helices, and one on an α3 helix. Table 3 compares these peptide sequences with those at the same amino acid sequence positions of HLA-F and HLA-G, there being no matching sequences for HLA-A, -B, or -Cw. The amino acids on the α1 and α2 helices of HLA-E—respectively, (R65D69) and (S152)—are essential for recognition by the inhibitory receptors CD94 and NKG2A present on the cell surface of CD8+ T cells and NK cells. Figure 4 shows the structure of HLA-E, pinpointing the precise location of the two peptide sequences, one on an α1 helix (65RSARDT70) and the other on an α2 helix (154AESADNSKQES144), which shows that these sites are exposed on the cell surface of HLA-E (with or without exogenous peptides) and are involved in direct binding to the amino acid sequences of CD94 and NKG2A. The unique HLA-E sequences of amino acids were synthesized and tested to determine how well the anti-HLA-E monospecific MAb TFL-033 was inhibited. Figure 5 shows the results: the binding of TFL-033 to HLA-ER107 was inhibited in a dose-dependent manner. Of the two peptides tested, 154AESADNSKQES144 showed better inhibition. However, one cannot rule out the possibility that the MAb bound to both α1 and α2 peptides since the location of the amino acid sequences appear adjacent in the surface view of the structure (Fig. 4).

Structure of HLA-E, showing the amino acid sequences on the α1 (65RSARDTAQIFRVNLR79) and α2 (162DELYARQHEAESADNSKSQES151) helices, which are specific for HLA-E and are not found in other classical HLA-Ia (HLA-A, -B, and -Cw) or non-classical HLA-Ib (HLA-F and -G) molecules, and therefore constitute the specific binding site for recognition by the monospecific anti-HLA Abs. The amino acids in the α1 (R65D69Q72I73R75V76R79) and α2 (E166D162A158H155E154S151) helices of HLA-E bind to amino acids on CD94 (Q112Q113F114L162D163E164N170) and NKG2A (R137P171S172K199Q212K217).(24)
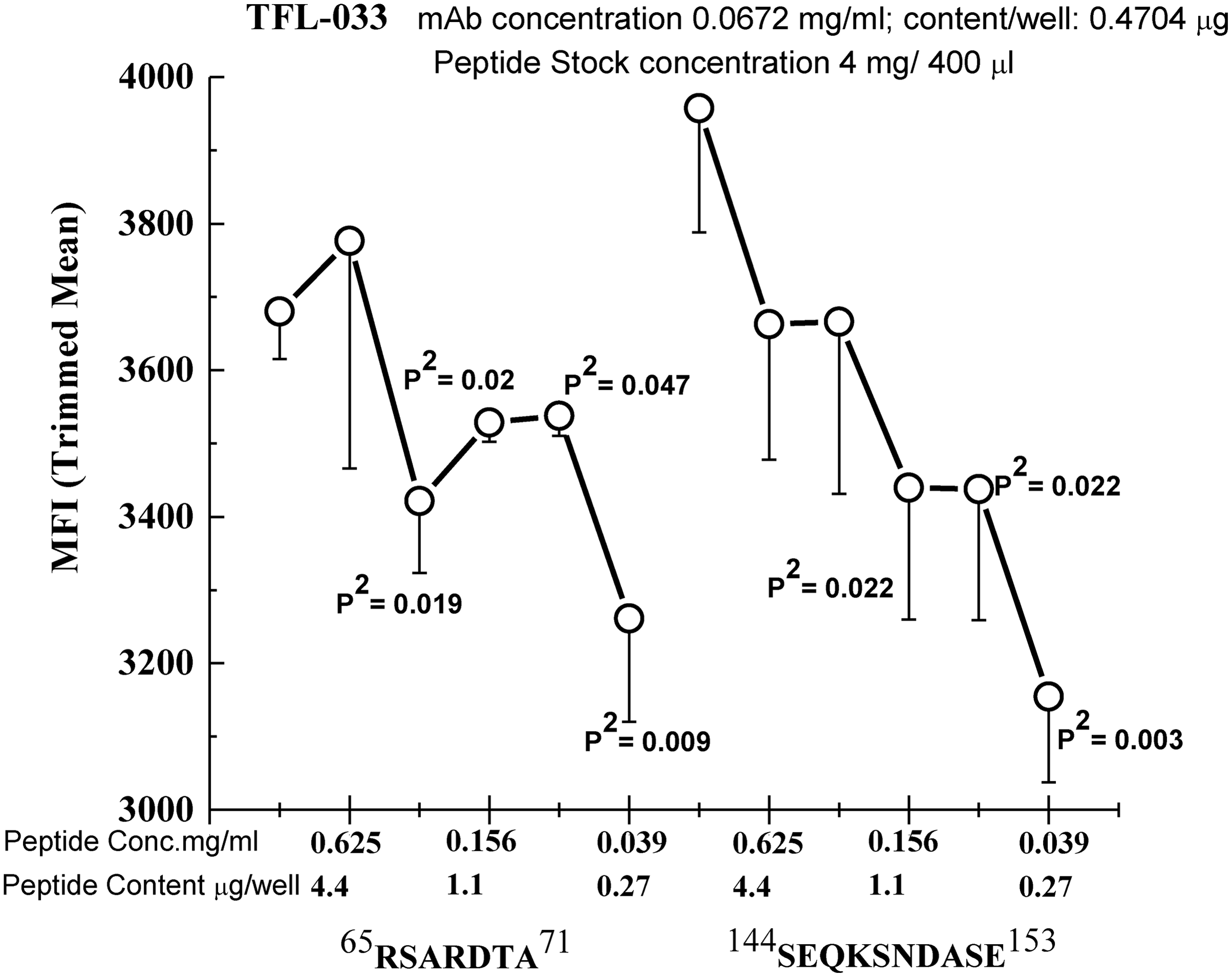
Dosimetric inhibition of purified culture supernatants of TFL-033 with two HLA-E-restricted peptides, 65RSARDTA71 and 143SEQKSNDASE152, at concentrations ranging from 4.4 to 0.27 μg/well. Although both peptides showed inhibition, the α2 helical peptide SEQKSNDASE showed better dosimetric inhibition than the other peptide.
Immunodiagnosis of HLA-E with mono- and polyspecific anti-HLA-E MAbs on HLA-E immunostaining
In this investigation, as was done previously,(25) HLA-E reactivity of the anti-HLA-E monospecific MAb TFL-033 was compared with that of the HLA-E- and HLA-Ia-reactive MAb MEM-E/02. We followed two sets of postulates as a guideline to understanding the rationale underlying the immunodiagnosis of TFL-033 compared with that of MEM-E/02:
• If a region of a tissue or tumor is positive to both MAbs, it implies reactivity of the MAbs to β2m-free HLA-E open conformers because MEM-E/02 reacts to the shared peptide sequences of HLA-E and HLA-Ia (117AYDGKDYLT125, 126LNEDLRSWTAV136, and/or 137DTAAQI 142).(27,28) Although TFL-033 is monospecific, it can still recognize the HLA-E-specific unique peptides in the α1 and α2 helices on the open conformers. • If a region of the tissue or tumor is TFL-033-positive and MEM-E/02-negative, it implies the absence of the binding site of MEM-E/02—the same shared peptide sequences of HLA-E and HLA-Ia as above (27,28,32,33)—which is possible when these sites remain cryptic due to the presence of β2m, confirming the presence of intact HLA-E with or without exogenous peptides. It is known that β2m is lost after shedding the exogenous peptide. Since the amino acid sequences in α1 and α2 helices are the binding sites of TFL-033, immunostaining confirms the presence of intact HLA-E.
These alternative immunostaining results were examined using human melanoma and diffuse gastric carcinoma microarrays.
Immunostaining of melanoma with MEM-E/02 versus monospecific MAbs
Figure 6A illustrates immunostaining of human melanoma microarrays with MEM-E/02 and with monospecific monoclonals in culture supernatants (s) including TFL-033s, -034s, -073s, and -145s, all of them with high isotype-specific MFI (>20 K when tested neat). The results were compared with stained sections that were not treated with primary MAbs: all monospecific MAbs and MEM-E/02 showed identical staining. While background staining was highly prevalent with MEM-E-02, immunostaining with TFL-MAbs was sharp and distinct. Since both MEM-E/02 and the monospecific TFL MAbs stained similar locations, we inferred that the MAbs were reacting to β2m-free HLA-E open conformers, as indicated above.
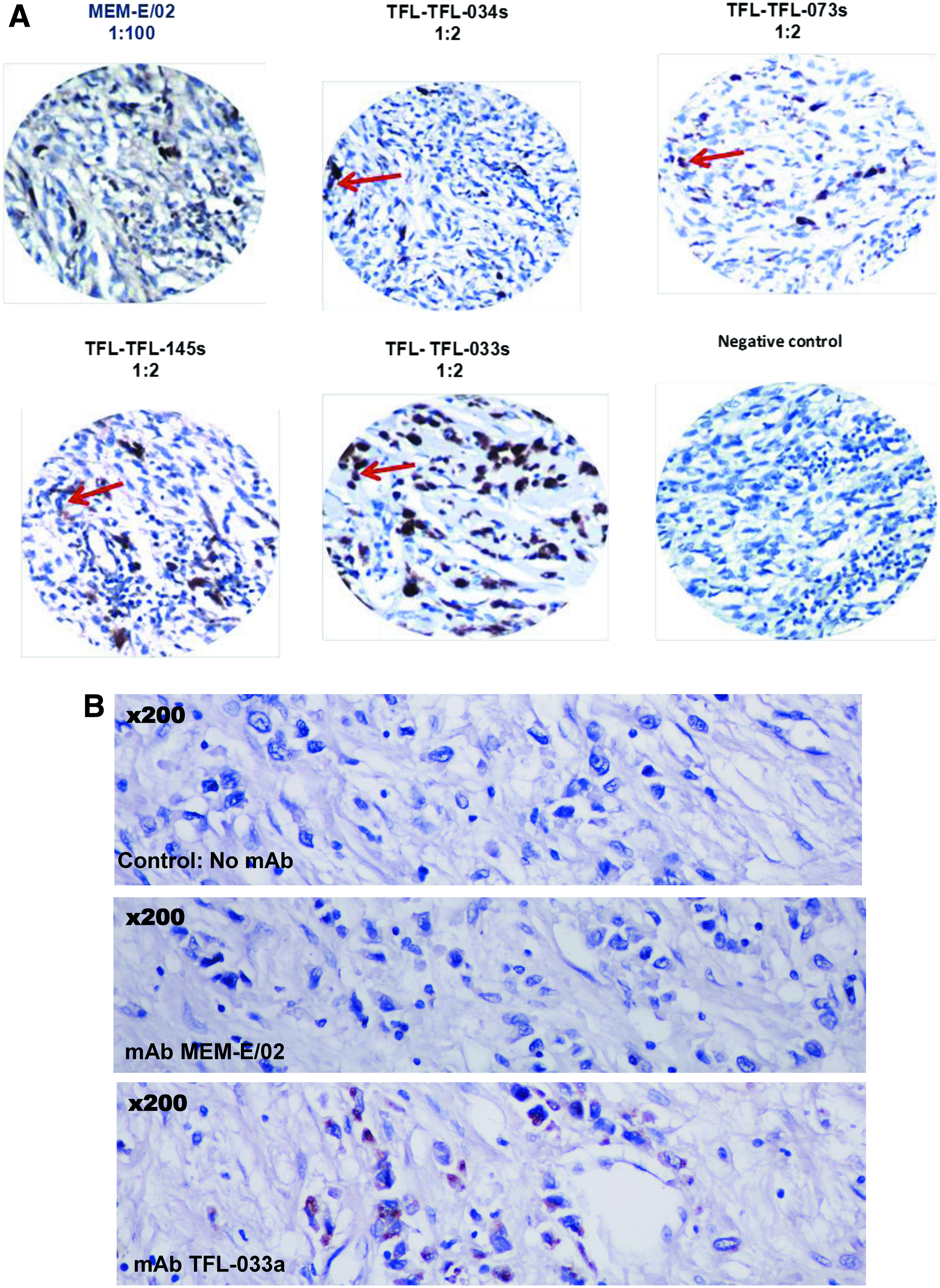
Immunolocalization of HLA-E in cancer tissues with culture supernatants (s) or ascites (a) of TFL monospecific MAbs compared with staining by MEM-E/02, an HLA-E MAb that shows cross-reactivity to HLA class Ia alleles. (
Immunostaining of diffuse carcinoma with MEM-E/02 versus TFL-033
Figure 6B illustrates immunostaining of human diffuse gastric carcinoma with MEM-E/02 and with the ascites of TFL-033. It is already known that both the incidence and staining intensity of TFL-033 is much better than MEM-E/02 for human diffuse gastric carcinoma.(25) The noteworthy observation here is that MEM-E/02 did not stain diffuse carcinoma while TFL-033 showed strong reactivity. The non-immunostaining by MEM-E/02 and positive reactivity of TFL-033 indicate the presence of intact HLA with β2m or clustering of cell surface over-expressed HLA-E masking the amino acid sequences shared by HLA-E and HLA class Ia alleles. The MAb MEM-E/02 failed to immunostain because the shared epitopes were cryptic. But since the amino acid sequences on the α1 and α2 helices were exposed and available, TFL-033 alone could recognize the epitopes and show positivity.
Monospecific HLA MAbs specifically augment the number of both untreated and PHA-treated CD4-/CD8+ T lymphoblasts
Table 4 presents data from the flow cytometry analysis of the effects of two dilutions of the monospecific MAbs TFL-033s, -034s, -073s, -074s, and -145s obtained from the serum-free culture supernatants on human T lymphocytes in vitro. The T lymphocytes were divided into three groups based on staining and size of cells and on T cell number (or events). Group 1 comprises resting CD4+ and CD8+ lymphocytes; group 2, resting CD4+ and CD8+ lymphocytes; and group 3, CD4+ and CD8+ lymphoblasts. The table documents the effects of MAbs on untreated (no PHA) and PHA-treated T lymphocytes in these categories of T cells: CD4+/CD8-, CD4-/CD8+, CD4+/CD8+, and CD4-/CD8-. The table compares the effect of MAbs on the number of untreated and PHA-treated cells with the number of cells when they were not treated with MAbs (control), showing the significant increase or decrease in the number of cells for both experimental and control groups for each MAb at each dilution.
MAbs TFL-033s, TFL-034s, TFL-073s, TFL-074s and TFL-145s.
All experiments were in triplicate. The T lymphocytes were obtained from a TFL staff member (J).
Figure 7 shows that monospecific MAbs mediated significant upregulation of CD4-/CD8+ T lymphoblasts. There was a significant increase in the number of CD4-/CD8+ T lymphoblasts among the PHA-treated T lymphoblasts under the influence of TFL-033s at 1:30 and 1:150, and TFL-034s at 1:50 whereas the number of PHA-untreated T lymphoblasts increased for almost all MAbs (TFL-033s at 1/30 and 1/150, TFL-034s at 1/10 and 1/50, TFL-073s at 1/50, TFL-074s at 1/10, and TFL-145s at 1/20). The increase in PHA-untreated T lymphoblasts clarifies the functional potential of HLA-E monospecific MAbs in augmenting CD4-/CD8+ T lymphoblasts.

Increase in the number of cells and percentage of non-activated and PHA-activated CD4-/CD8+ T lymphoblasts (group 3; Table 4) 72 h after exposure to two dilutions of purified supernatants of TFL-033, -034, -073, and -145 in vitro, in serum-free culture.
Discussion
Characteristics of monospecific anti-HLA-E MAbs
Twelve percent of the clones produced after immunization with heavy chain of rHLA-EG107 or rHLA-ER107 secreted monospecific anti-HLA-E MAbs. The number of clones are higher after immunization with rHLA-ER107 than with HLA-EG107, possibly due to the rarity of arginine at position 107; R107 occurs only in HLA-A*01010102N and -B*1530 in contrast to glycine (G) at 107, which is common to HLA-Ia, HLA-F, and HLA-G (Table 3). The rarity of R107 may account for the better immunogenicity of HLA-ER107 in both mice and humans, although HLA-EG107 and HLA-ER107 alleles commonly coexist in different human ethnicities.(52) The prevalence of anti-HLA-E Abs found in normal healthy non-alloimmunized humans,(47) in cancer patients,(48) and in allograft recipients(49) could be due to the dominant expression(52) and immunogenicity of HLA-ER107.
Recognition of HLA-E-specific amino acid sequences (Table 3) by MAbs is a defining characteristic of the monospecificity of anti-HLA-E MAbs. The dosimetric inhibition of HLA-E binding of the monospecific MAb TFL-033 by HLA-E-restricted amino acid sequences (65RSARDT70 and 154AESADNSKQES144) (Fig. 5) confirms that the recognition sites of the MAb TFL-033 are the epitopes expressed on the α1 and α2 helices. The monospecific MAbs are probably capable of recognizing discontinuous sequences on both of the helices or a continuous sequence on one of them. For studies involving the immunodiagnosis of HLA-E expression in cancer cells—and for testing the immunomodulatory potential of the anti-HLA-E monospecific MAbs—we used only the monospecific MAbs generated after immunization with HLA-ER107: TFL-033, −034, −073, −074, and −145. We speculate, but have not proven, that the results would be the same with monospecific MAbs generated after immunization with HLA-EG107.
Immunodiagnostic potential of monospecific anti-HLA-E MAbs
In an earlier report, we confirmed the reactivity of the monospecific MAb TFL-033 on gastic cancer cell lines (AGS, primary adenocarcinoma cell line, negative for HLA-E by both MEM-E/02 and monospecific MAb TFL-003; SNU-1, peritoneal metastatic cell lines; KATO III, lymph node metastatic cell lines; and NCI-N87, liver metastatic cell lines), expressing different levels of HLA-E and upregulated HLA-E after exposure to IFNγ and TNFα. MEM-E=02 stained HLA-E post-IFNγ in SNU-1, KATO-III, and NCI-N87 cell lines, whereas TFL-033 stained KATO-III and NCI-N87. The positive signals of KATO-III and NCI-N87 were amplified dosimetrically with IFNγ, up to 20% (see Fig. 3 in ref 25) but were not amplified using either TNFa (6%) or IL-6 (0.6%). These observations provide better validation and appraisal of the monospecific MAb TFL-033.(25)
The use of HLA-polyreactive commercial anti-HLA-E MAbs for immunodiagnosis of HLA-E is untenable and misleading because of their cross-reactivity with HLA-Ia alleles. In contradistinction, monospecific anti-HLA-E MAbs are reliable for immunodiagnosis of HLA-E expression on cancer tissues. However, it is important to note that since mono- and polyspecific anti-HLA-E MAbs differ in their recognition site, both kinds of MAbs can be used to distinguish expression of intact HLA-E (associated with β2m) from open conformers of HLA-E (β2m-free), as reported in this study and in an earlier report.(25)
Indeed, Sasaki and colleagues applied this strategy for immunodiagnosis of the HLA-E expression on gastric cancer tissues.(25) It was noted that the incidence and intensity of TFL-033 staining of the cell surface in the early stages, and with poorly or undifferentiated and non-nodal lesions, and in diffuse gastric carcinoma, was greater than the incidence and intensity of staining by MEM-E/02; however, MEM-E/02 did stain terminal stages, gastric adenocarcinoma, and lymph node metastatic lesions more intensely than it stained diffuse carcinoma. In the present study, there was immunostaining of human diffuse gastric carcinoma with monospecific MAb TFL-033 (Fig. 6B) but not with polyspecific MAb MEM-E/02. The failure of MEM-E/02 to immunostain diffuse gastric carcinoma—while TFL-033 did—demonstrates the expression of intact HLA-E associated with β2m in diffuse carcinoma. The immunostaining of human melanoma tumor tissues by MEM-E/02 shows the presence of HLA-E open conformers in human melanoma. All the monospecific MAbs (including TFL-033) used to immunostain human melanoma (Fig. 6A) also stained tissue sections of melanoma in a similar fashion. The differences in the location of the peptides recognized by MEM-E/02 and TFL-033, and the simultaneous exposure of the different peptide epitopes in the open conformer of HLA-E, favor the simultaneous use of these two kinds of MAbs (mono- and polyspecific) to differentiate intact HLA-E from β2m-free HLA-E.
A number of reports (cited above in the Introduction) document the presence or overexpression of HLA-E in different cancer tissues, the underlying studies using commercially available polyspecific anti-HLA-E MAbs for immunolocalization of HLA-E. Unfortunately, these reports do not provide authentic and specific immunodiagnosis of HLA-E because the polyspecific HLA-E MAbs used in these studies (MEM-E/02, -E/06, -E/07, -E/08, or 3D12) react with shared epitopes located on the heavy chain of HLA-E (cryptic in the presence of β2m) or the open conformers of both HLA-Ia and HLA-Ib alleles. Figures 8A and 8B compare the HLA-E specific peptide sequences recognized by monospecific MAbs (e.g., TFL-033) with the peptide sequences shared among all HLA class I alleles, including HLA-E. Figure 8A shows the specific amino acids (shown in yellow) located in the α1 and α2 helices of HLA-E recognized by monospecific MAbs (e.g., TFL-033), which are accessible to the Abs in the presence as well as in the absence of β2-microglobulin. Figure 8B shows the commonly shared but cryptic peptide sequences of HLA-E (shown in yellow) in the vicinity of β2-microglobulin recognized by polyspecific MAbs (e.g., TFL-006 and TFL-007), when HLA is free of β2-microglobulin or otherwise known as open conformers. Although some reports(1–3) document that HLA class-Ia alleles (HLA-A, -B, and -Cw) are lost or downregulated, such a generalization cannot be extended to all tumors. In support of that contention, a report(52) analyzing a microarray consisting of cancer tissues from 285 stage I-IV colon cancer patients showed that HLA-Ia and HLA-E were expressed, respectively, by 79.8% and 75.8% of the tissues while loss and downregulation of HLA-Ia and HLA-E were observed in only 4.5% and 15.7%, respectively. In addition, while the loss HLA-Ia does occur in different human cancer types, it occurs only to a limited extent.

Structure of intact HLA-E showing peptide sequences recognized by the anti-HLA-E MAbs. (
Immunotherapeutic potential of monospecific anti-HLA-E MAbs
Our results further show that in addition to the particular potential of monospecific anti-HLA-E MAbs to block ligands of inhibitory receptors on the α1 and α2 helices of HLA-E, these MAbs are also capable of triggering the proliferation of both non-activated and activated CD8+ T lymphocytes, which include both CD3+/CD8+ cytotoxic T cells and CD3-/CD8+ NK cells or NKT cells. This observation is especially striking because a significant increase in the number of CD8+ T lymphoblasts was observed even in the absence of an activating co-stimulant (PHA). This unique property of augmenting the number of CD8+ T lymphocytes is an added benefit and a further indication for the use of monospecific anti-HLA-E MAbs in cancer treatment.
These results document that the monospecific TFL MAbs are indeed augmenting the number of CD8+ cells, suggesting that these TFL MAbs are binding to HLA-E molecules expressed on the CD8+ T and NKT cells. HLA-E is a component of unspecified HLA class I antigens earlier identified at low levels on inactivated CD8+ cells and upregulated(54,55) or heavily clustered(56) in activated (by PHA or IFNγ) CD8+ cells.
In general, blastogenic transformation and proliferation result in transitory cell surface expression of several molecules including HLA class I and II molecules.(43) It is interesting to note that β2m-free heavy chains of HLA class I, called “open conformers” are also overexpressed and even clustered heavily on the activated T cells.(54–56) Several reports have documented the elongation of the cytoplasmic tail of the HLA-I open conformers, and the exposure of otherwise cryptic tyrosine 320(57,58) and serine 335(59) with a provision for phosphorylation. We hypothesize that the binding of the monospecific anti-HLA-E MAb onto α1 and α2 helices of the overexpressed and clustered open conformers of HLA-E in activated normal human CD8+ T and NKT cells may induce phosphorylation, promoting proliferation of both non-activated as well as PHA-activated CD8+ T and NKT cells at different levels, as presented in a model (Fig. 9). This model takes into consideration the report of Matko and colleagues(56) on the clustering of HLA-I molecules on the cell membrane of activated (either by PHA or IFNγ) CD8+ T lymphocytes. Since NKT cells are devoid of CD3 molecules, we envision that the TFL-MAb mediated activation of CD8+ NKT cells may be independent of CD3 molecules or may involve different cell surface receptors.
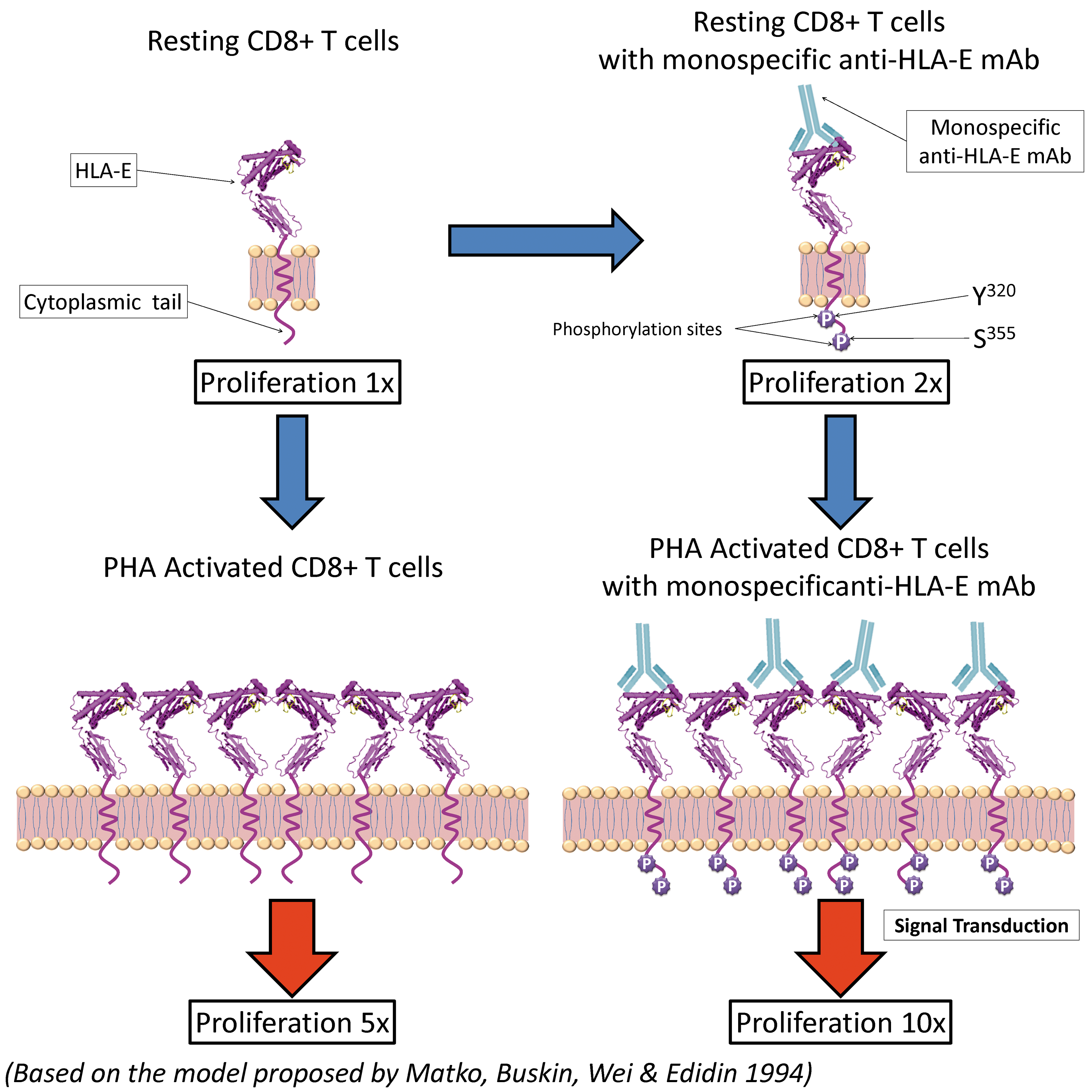
A model based on a previous report(56) illustrating the possible mechanism underlying proliferation non-activated and phytohaemagglutinin (PHA)-activated CD8+ T lymphocytes, which include cytotoxic T cells (CD3+) as well as NKT cells (CD3-) mediated by monospecific anti-human leucocyte antigen (HLA)-E monoclonal antibodies (Terasaki Foundation Laboratory) (TFL-033, TFL-034, TFL-073, TFL-145). Inactive (resting) CD8+ T cells express β2-microglobulin-free HLA-E or the open conformers. Their proliferation level is shown to be minimal (1x). Resting CD8+ T cells are activated by monospecific anti-HLA-E MAbs, which bind to α1 and α2 helices expressing HLA-E specific amino acid sequences (see Fig. 4), promoting further proliferations (2x) by phosphorylation of the tyrosyl and serine residues of the elongated cytoplasmic tail of the HLA-E open conformer. Elongation of the cytoplasmic tails of open conformers that results in exposure of cryptic tyrosine (Tyr320) and serine (Ser355) is well known.(43) The open conformer is situated on the lipid raft (pink zone) of the bi-layered lipid membrane in the non-phosphorylated non-activated CD8+ T cells. Binding of monospecific anti-HLA-E MAb by binding to the HLA-E open conformers may induce lymphocyte-specific protein tyrosine kinase (LCK) to phosphorylate the tyrosul residue located on the cytoplasmic tail of HLA-E. Activation may result in enhanced proliferation. The figure illustrates that it may be the binding of anti-HLA-E monospecific MAbs such as TFL-033 that initiates the phosphorylation leading to signal transduction. A model proposed by Matko and colleagues(56) recognizes the clustering of HLA-I molecules overexpressed on activated (by PHA or IFNγ) CD8+ T cell in contrast to inactivated CD8+ cells. Data presented in Figure 7 show increased proliferation of PHA activated CD8+ T cells upon exposure to anti-HLA-E monospecific MAbs listed above. Not all anti-HLA-E MAbs are capable of inducing proliferation of activated CD8+ T cells (see Table 4). If proliferation is 5x after PHA activation, exposure to HLA-E monospecific MAbs increases the proliferation to 10x. Though x values are approximate, the data presented in Figure 7 clearly indicates that it may differ with different HLA-E monospecific MAbs. We have not shown phosphorylation of PHA activated CD8+ T cells because the PHA activation is mediated through other receptors like CD3. Such activation leads to overexpression of open conformers of HLA-E and clustering of the open conformers, as proposed by Matko and colleagues.(56) Such clustering may facilitate better binding of anti-HLA-E monospecific MAbs and increased proliferation.
HLA-E may transmit inhibitory signals mediated by CD94/NKG2A present on CD8+ T cells and NKT cells. The siRNA-mediated silencing of HLA-E or blocking of CD94/NKG2A enabled lysis of 51Cr-labeled tumor cells.(36) Since one or more of the amino acids α1 and α2 recognized by the anti-HLA-E MAbs serve as the recognition and binding sites of the inhibitory receptors of NK and CD8+ αβ and γδ T cells, these monospecific anti-HLA-E MAbs could serve as immunotherapeutic agents with the potential of blocking both HLA-E expressed on the tumor cells and their interaction with the inhibitory receptors of the CD8+ cytotoxic as well as NKT cells that have anti-tumor potential, but remain paralyzed due to interaction of tumor associated HLA-E with inhibitory receptors on CD8+ T cells. Such blocking may restore the cytotoxic capabilities of CTLs and CD8+ NK cells, which would otherwise remain inactivated by HLA-E on the tumor cell surface. Moreover, blocking HLA-E could be a better strategy than blocking CD94/NKG2A receptors since they continuously recycle from the cell surface through the endosomal compartments and back again to the cell surface, a process that requires energy and the cytoskeleton.(51)
Conclusion
In sum, it may be stated that the monospecific anti-HLA-E MAbs reported here are superior to commercially available anti-HLA-E MAbs in their ability to identify and specifically localize HLA-E expression on the cell surface of tumor tissues. In addition, they can serve a dual role in combating cancer by blocking HLA-E inhibitory receptor interaction and simultaneously inducing proliferation of CD8+ CTL in the presence or absence of co-stimulation.
Footnotes
Acknowledgments
We thank Dr. Cecilia M. Freitas and Mr. Satoru Kawakita at TFL for immunostaining and photographic documentation of melanoma and gastric carcinoma microarrays.
Author Disclosure Statement
This research project was supported entirely by grants from the Terasaki Family Foundation. There is no conflict of interest for any of the authors. However, a patent on monospecific anti-HLA-E MAbs filed by Ravindranath and Terasaki in 2013 contains the salient findings reported in this manuscript.