
Abstract
Four hybridoma cell lines (C4B, 10C2G5, 6C5F4C7, 2D10G11) were adapted to grow in serum-free conditions. Cell proliferation, viability, and antibody production in Nutridoma SP and Ex-Cell HSF 610 media were evaluated and results compared with those obtained using DMEM containing 10% fetal bovine serum (control medium). Clone C4B showed the best IgG productivity in control medium, but viability and number of cells per milliliter were similar for the three media. For clone 10C2G5, the highest values of cell viability were obtained with both control medium and Nutridoma SP; no significant differences in IgG yields and number of cells/mL were observed among the three media. Clone 6C5F4C7 provided no significant differences when grown in the two serum-free media and in control medium. Clone 2D10G11 showed the best IgG productivity in control medium and in Ex-Cell HSF 610, even if Ex-Cell HSF 610 provided the lowest number of cells/mL; no significant differences among the three media were obtained for viability. Purified antibodies produced from each hybridoma cell line grown in serum-free media showed a higher degree of purity than those produced by the same cell lines grown in control medium. Purified MAbs were also titrated by ELISA to test MAb-antigen affinity.
Introduction
T
The aim of our study was to assess cell growth and antibody production of four different hybridoma cell lines (C4B, 10C2G5, 6C5F4C7, 2D10G11) cultured in two serum-free media, Ex-Cell 610 HSF and Nutridoma SP, for their use in lab-scale production of MAbs.
Materials and Methods
Hybridoma cell lines
Four hybridoma cell lines, produced at the Istituto Zooprofilattico Sperimentale dell'Abruzzo e del Molise “G. Caporale” (Teramo, Italy) by fusion between SP2/0-Ag14 myeloma cells and murine splenocytes,(15,16) were used in this study. Clones C4B, 10C2G5, and 2D10G11 produce IgG1 anti k antibodies against, respectively, bovine IgG, swine IgG, and VP7 of African horse sickness virus (AHSV); clone 6C5F4C7 produces IgG2a anti k antibodies against VP7 of Bluetongue virus (BTV).
Cell line adaptation and culture conditions
After thawing, each hybridoma cell line was initially grown in DMEM containing 10% FBS (v/v) (Sigma, St. Louis, MO), 2 mM glutamine (Sigma), 100x penicillin-streptomycin-amphotericin (Sigma), 50 mg/mL gentamicin (Sigma), and 10000 IU/mL nystatin (Sigma). Then the clones were adapted to grow in the presence of decreasing concentrations of FBS (5%, 2.5%, 1%),(2) initially in 25 cm2 flasks and subsequently in flasks of increasing volume (75-175-525 cm2). During the adaptation phase, the cells were cultured at +37°C in a 90% humidified atmosphere with 5% CO2 and were maintained at the same concentration of serum for at least two passages. At the end of adaptation phase, all the hybridoma cell lines were grown in the serum-free media Ex-cell HSF 610 (Sigma) and Nutridoma SP (Roche, Mannheim, Germany). The cultivation of the hybridomas in DMEM containing 10% FBS (v/v) (DMEM) was used as a control. The clones cultivated in the medium Ex-cell HSF 610 were grown under the conditions mentioned above; cells were initially seeded at a density of 3–5×105 viable cells/mL, while after complete adaptation, cells were seeded at a density of 2×105 viable cells/mL. The clones cultivated in the Nutridoma SP medium were again grown at +37°C in a 90% humidified atmosphere with 7.5% CO2, but cells were seeded at a density of 1–2×105 viable cells/mL. The number of cells/mL and the percentage of cell viability were determined at each cell passage by an automatic cell counter (Countess® Automated Cell Counter, Invitrogen, Carlsbad, CA). Adherent cells cultivated in DMEM were passed for expansion when they reached 80–90% confluence; non-adherent cells cultivated in serum-free media were sub-cultured two to three times a week in order to maintain the recommended sub-culture seeding for each medium (2×105/mL for Ex-Cell HSF 610; 1–2×105/mL for Nutridoma SP), according to the medium data sheet.
Hybridoma cell line cultivation in the two serum-free media and in DMEM was carried out in triplicate; for each assay 1 L of supernatant was produced.
Antibody purification
Supernatants from each culture were centrifuged twice at 400 g for 10 min at +4°C. In order to evaluate the IgG production for each clone grown in the two serum-free media and in the control medium, supernatants were filtered through a 0.22 μm filter and loaded onto a HiTrap rProtein A FF column (GE Healthcare, Uppsala, Sweden), according to the manufacturer's instructions. Purified IgG were concentrated using 100 kDa cut-off centrifugal filters (Millipore, Billerica, MA) and resuspended in PBS 0.01 M (pH 7.2). The protein concentration of purified MAbs was determined by spectrophotometry (Absorbance at 280 nm/IgG molar extinction coefficient).
To compare the yields of IgG produced by the MAbs in both serum-free media and control medium, the purification of bovine IgG, present in a liter of DMEM, was carried out by affinity chromatography, as indicated above.
Statistical analysis
The number of cells/mL, their viability (%), and IgG production were analyzed with descriptive statistics, by calculation of means, standard deviations, and confidence intervals at 95% probability. When the 95% confidence intervals of the mean did not overlap, the differences were considered statistically significant.(17)
Antibody titration by indirect ELISA
Purified MAbs were titrated in indirect ELISA using Costar High Binding microplates (Corning, New York, NY) coated overnight with 10 μg/mL of the related antigen (purified bovine and swine IgG, AHSV, and BTV whole virus) diluted in 0.05 M carbonate-bicarbonate buffer (pH 9.6), and blocked with 1% yeast extract for 1 h at room temperature (RT). One hundred μL/well of purified MAbs, serially diluted 2-fold from 100 μg IgG/mL to 0.003 μg IgG/mL, were incubated for 1 h at +37°C. After washing with PBS containing 0.05% Tween-20 (PBST), 100 μL/well of ECL anti-mouse-IgG-HRP (GE Healthcare) diluted 1:8000 were added, plates were left for 30 min at RT and then washed with PBST. After addition of 100 μL/well of 3,3', 5,5′-tetramethylbenzidine substrate (Sigma), plates were again incubated at RT for 30 min. The reaction was stopped by addition of 50 μL/well of 0.5 N H2SO4. Optical density was measured at 450 nm (OD450nm).
SDS-PAGE
The degree of purity of purified MAbs produced in serum-free media and control medium was evaluated by SDS-PAGE. Proteins in media and MAbs were separated by electrophoresis in denaturing conditions using NuPage® 12% Bis-Tris pre-cast gels (Invitrogen) at a constant voltage of 200 V. At the end of the run, gels were incubated overnight at RT with SimplyBlue Safestain (Invitrogen). Images were acquired with Chemidoc MP (Bio-Rad, Hercules, CA) using the Quantity One Quantitation Software (Bio-Rad).
Results
The amount of bovine IgG reacting with protein A (especially IgG2),(18) present in 1 L DMEM, was evaluated by affinity chromatography. The quantity of IgG obtained was 3.7 mg.
Comparative results of the performance of the four clones in the three different media are provided in Table 1. Clone C4B showed no differences in cell viability and number of cells/mL when cultured in control medium and in the two serum-free media.The best IgG yield was obtained in the control medium (54.0±12.6 mg), even taking into account the bovine IgG contamination due to the use of FBS (estimated at 3.7 mg/L of medium). No difference in the IgG yield among the serum-free media was recorded (13.8±3.5 mg for Nutridoma SP and 17.2±3.1 mg for Ex-Cell HSF 610; Fig 1).

Growth of clone C4B in DMEM 10% FBS and in serum-free media: viability (
Values shown represent mean and confidence interval (α=95%). Trend of viability and cell growth are shown: ↔, stable trend;↑increase;↓decrease.
For clone 10C2G5, no difference was recorded among the three media tested in terms of number of cells/mL. Moreover, no difference was obtained in viability between DMEM and Nutridoma SP and between DMEM and Ex-Cell HSF 610, but significant differences were obtained between the two serum-free media. Considering the bovine IgG contamination of 3.7 mg due to FBS, there was no difference among the IgG yields in the three media (5.8±0.5 mg for DMEM, 1.9±0.3 mg for Ex-Cell HSF 610, 3.0±0.1 mg for Nutridoma SP) (Fig. 2).

Growth of clone 10C2G5 in DMEM 10% FBS and in serum-free media: viability (
Clone 6C5F4C7 showed no differences in terms of viability and number of cells/mL for the three media. Even considering the estimated bovine IgG contamination of MAb produced in DMEM medium, the highest IgG yields were provided by DMEM and Ex-Cell HSF 610 (12.0±3.1 mg for DMEM, 9.3±0.3 mg for Ex-Cell HSF 610). With Nutridoma SP, the lowest IgG yield (1.9±0.2) was observed (Fig. 3).
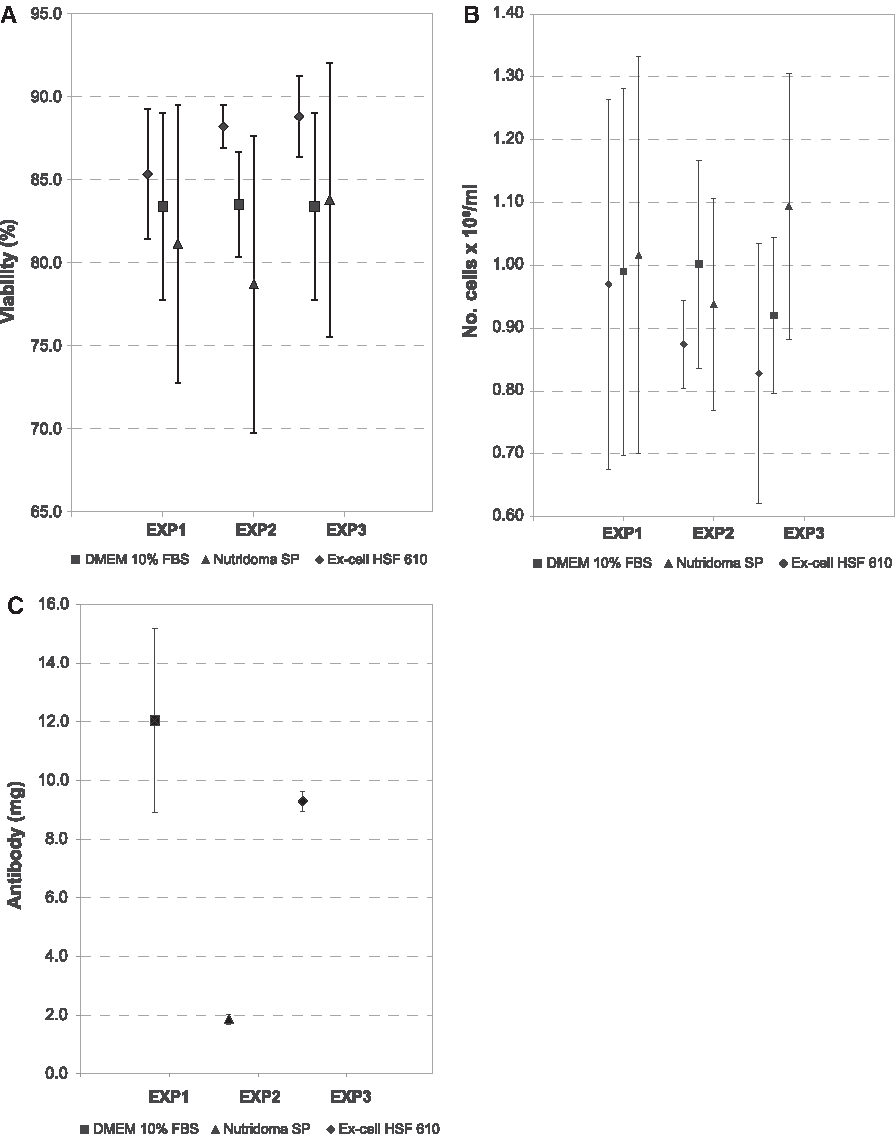
Growth of clone 6C5F4C7 in DMEM 10% FBS and in serum-free media: viability (
Finally, for clone 2D10G11, no difference was obtained in terms of viability among the three media tested. No difference was also obtained in number of cells/mL between Ex-Cell HSF 610 and Nutridoma SP and between DMEM and Nutridoma SP, but significant differences were obtained between DMEM and Ex-Cell HSF 610. The highest IgG yields were provided by DMEM and Ex-Cell HSF 610 (24.5±1.1 mg for DMEM; 18.9±1.7 mg for Ex-Cell HSF 610); considering the bovine IgG contamination of purified MAbs grown in DMEM, differences in IgG yields between DMEM and Ex-Cell HSF 610 could be considered not significant (Fig. 4).
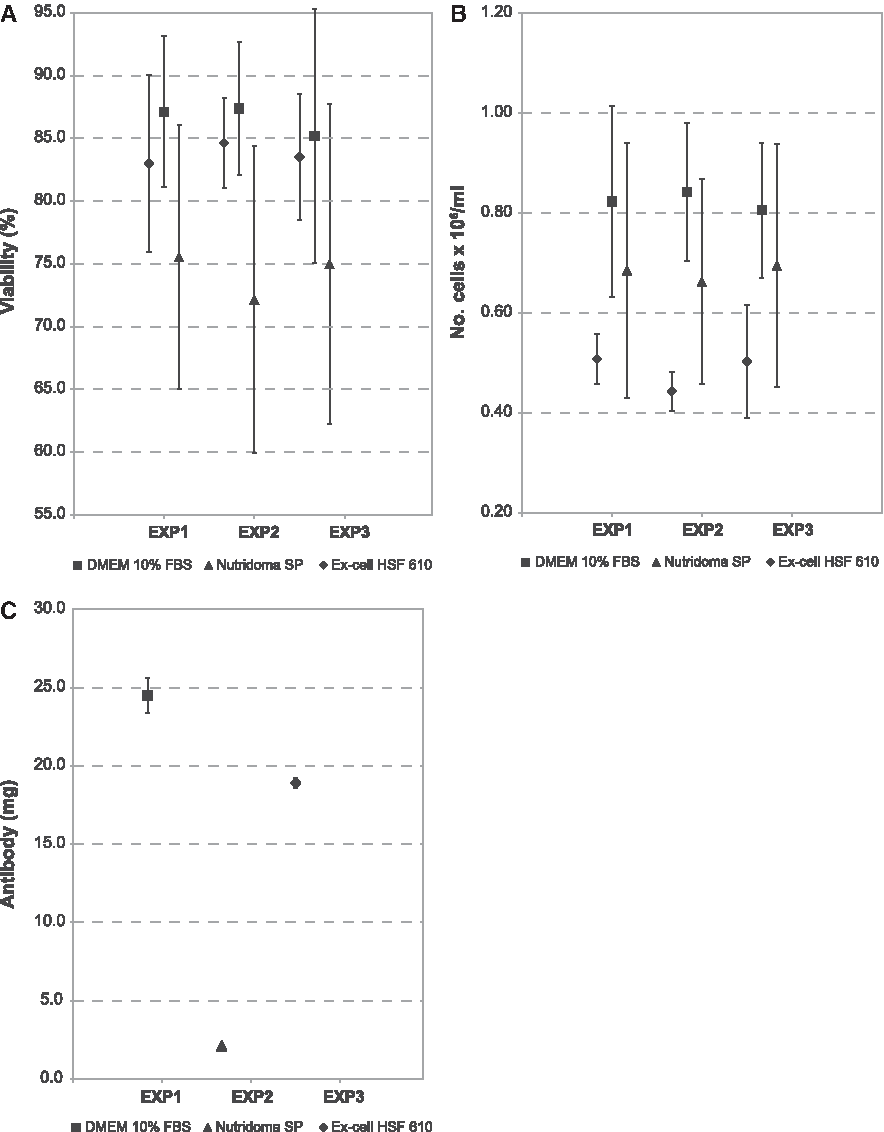
Growth of clone 2D10G11 in DMEM 10% FBS and in serum-free media: viability (
To assess their performance, the purified MAbs were diluted and titrated by ELISA. From the data obtained, it was found that at the higher concentrations (100–12.5 μg/mL), MAb C4B had a similar trend irrespective of the medium used, whereas at lower concentrations (≤6.25 μg/mL), the OD of MAb from serum-free media decreases more rapidily than that of the same MAb in DMEM (Fig. 5).
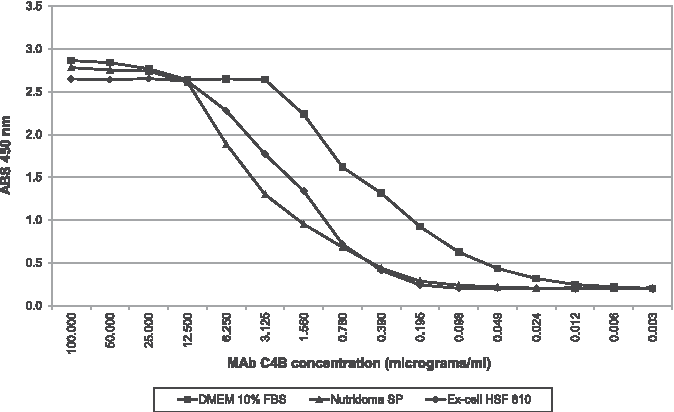
Indirect ELISA: titration of MAb C4B. Solid diamonds denote Ex-Cell HSF 610 results; solid squares denote DMEM 10% FBS results; solid triangles denote Nutridoma SP results.
The curves relating to the MAb 10C2G5 produced in DMEM and Ex-cell 610 HSF showed, at the highest concentrations (100–6.25 μg/mL), similar trends, while below 3.125 μg/mL the OD of MAb produced in DMEM decreased more rapidly. The MAb produced in Nutridoma SP medium showed the lowest OD at all IgG concentrations tested (Fig. 6).

Indirect ELISA: titration of MAb 10C2G5. Solid diamonds denote Ex-Cell HSF 610 results; solid squares denote DMEM 10% FBS results; solid triangles denote Nutridoma SP results.
At concentrations between 100 and 0.78 μg/mL, the curves of MAb 6C5F4C7 produced in the three media overlap; below the latter concentration, the curve of MAb produced in DMEM decreased more rapidly than that of the other two (Fig. 7).
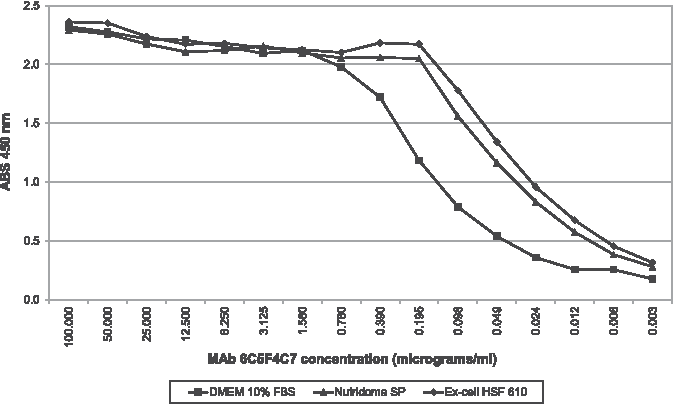
Indirect ELISA: titration of MAb 6C5F4C7. Solid diamonds denote Ex-Cell HSF 610 results; solid squares denote DMEM 10% FBS results; solid triangles denote Nutridoma SP results.
MAb 2D10G11 from Nutridoma SP showed the highest OD, while the products obtained with the other two media had a similar trend (Fig. 8). As expected, SDS-PAGE analysis of the purified MAbs produced in presence of serum-free media showed a higher degree of purity than those produced in the control medium (Fig. 9).

Indirect ELISA: titration of MAb 2D10G11. Solid diamonds denote Ex-Cell HSF 610 results; solid squares denote DMEM 10% FBS results; solid triangles denote Nutridoma SP results.

SDS-PAGE and Comassie stain of culture media and purified MAbs. Nutridoma SP (1); Ex-Cell HSF 610 (2); DMEM 10% FBS (3); MAb 10C2G5 produced in Nutridoma SP (4), Ex-Cell HSF 610 (5), DMEM 10% FBS (6); MAb 6C5F4C7 produced in Nutridoma SP (7), Ex-Cell HSF 610 (8), DMEM 10% FBS (9); MAb 2D10G11 produced in Nutridoma SP (10), Ex-Cell HSF 610 (11), DMEM 10% FBS (12); MAb C4B produced in Nutridoma SP (13), Ex-Cell HSF 610 (14), DMEM 10% FBS (15).
Discussion
The aim of the present study was an assessment of the use of serum-free media for the production of four hybridoma cell lines. MAbs were produced in liter volume and all tests were carried out in triplicate. Data obtained were statistically analyzed.
Speed of adaptation to serum-free growth conditions may be clone-dependent; some of them require a long period of adaptation in order to obtain stable conditions of growth. As already described,(19–22) production of IgG may be dependent on the culture medium used and on the growth conditions; for some clones IgG production is higher in media containing serum. Serum appears to have a regulatory as well as a stimulative effect on the antibody productivity(9) or could act as a cell protector from stresses.(22,23) The four clones under study (C4B, 10C2G5, 6C5F4C7, 2D10G11) were successfully grown in Nutridoma SP and Ex-Cell 610 HSF serum-free media. In terms of cell viability and cell number, clones C4B and 6C5F4C7 did not show differences when cultured in serum-free media and in DMEM; for clone 10C2G5, there was no difference in terms of number of cells among the three media tested, but a higher viability was obtained with Nutridoma SP compared with Ex-Cell 610 HSF. Viability of clone 2D10G11 obtained for DMEM and for the two serum-free media did not show differences, but DMEM provided a higher number of cells compared with Ex-Cell 610 HSF.
Clone 10C2G5 did not show differences in IgG yields when cultivated in any of the three media. Clone C4B showed a significantly higher yield with DMEM, while clones 6C5F4C7 and 2D10G11 showed similar yields both in DMEM and Ex-Cell 610 HSF; the yield in these two media is significantly higher than the yield in Nutridoma SP.
Regarding the purified MAbs, indirect ELISA titration results were variable. Clone 6C5F4C7 provided higher titers with both serum-free media, clone 2D10G11 with Nutridoma SP and clone 10C2G5 with Ex-Cell 610 HSF, whereas C4B with DMEM. One hypothesis that could explain these results is that the binding affinity of IgG for the specific antigen is influenced by the culture medium used. Some investigators have already shown that the type of medium used for adaptation and production of MAbs have not only effects on the growth of hybridomas and on the yield in IgG, but may change other parameters, such as the glycosylation profile of immunoglobulins, responsible for the changes of the intrinsic properties and the stability of IgG, modifying some biological functions related to complement dependent cytotoxicity and antibody-dependent cell-mediated cytotoxicity.(21,24–26)
Further studies must be carried out on the clones analyzed in this study to assess the possible influence of culture media on the binding affinity of the antibodies to the specific antigen.
The cultivation of the hybridomas in media not enriched by serum has the advantage of providing MAbs with a high degree of purity. It is known that some IgG subclasses present in FBS bind to proteins A and G of affinity columns used for antibody purification and remain as contaminants after purification,(4,10,18,27,28) giving rise, in some cases, to non-specific binding with antigens or with blocking agents used in immunoassays, resulting in increased background signals and decrease in sensitivity and specificity. Non-specific serum antibodies can be removed from purified MAbs solutions by subsequent immunoadsorption; however, the process can lengthen the time of production and increase the costs.
For the future, to produce MAbs with a higher degree of purity for veterinary diagnostics, it will be necessary to identify the best serum-free medium for each line of hybridoma, in order to obtain adequate IgG yields and optimal diagnostic performances.
In conclusion the use of serum-free media is a valid solution for the production of MAbs, since they provide products characterized by a high degree of purity with consequent reduction of non-specific reactions in diagnostic tests. Serum-free media are also useful to reduce the use of animal-derived reagents in MAbs production.
Footnotes
Acknowledgments
The authors wish to thank Dr. Attilio Pini for a critical reading of the article and Mrs. Paola Di Giuseppe for image editing.
Author Disclosure Statement
The authors have no financial interests to disclose.