
Abstract
The variable regions of the heavy chain (VH) and light chain (VL) were amplified by RT-PCR from the hybridoma 1N8, which secretes the monoclonal antibody against CDV N protein (aa 277-471). The VL and VH amplicons were combined using SOE-PCR by a 12 amino acid flexible linker (SSGGGGSGGGGS), which produced the scFv gene (named scFv/1N8). After sequence analysis, the scFv/1N8 gene was cloned into the prokaryotic expression vector PET32a with a His-tag. The recombinant scFv/1N8 protein was successfully expressed in recombinant Escherichia coli by IPTG induction. Moreover, the binding activity and specificity of the scFv were determined by indirect ELISA (His-tag) and competitive ELISA. The recombinant scFv/1N8 protein reported here will provide some basis for further antiviral drug research based on the scFv molecule.
Introduction
C
Since CD was first reported and described in detail at the beginning of the 19th century by Jenner, many studies regarding CDV have been carried out for the prevention. However, in recent years CDV infections still exist(6) and cause frequent occurrences in many fur animal–raising provinces in China, leading to severe economic losses.
Canine distemper virus is a Morbillivirus of the family Paramyxoviridae; it is an enveloped, negative-sense, single-stranded RNA virus that contains six structural proteins, the nucleoprotein (N), the phospho- (P), the large (L), the matrix- (M), the hemagglutinin (H), and the fusion (F) protein. Additional accessory genes, the C and V proteins, are found mostly as extra-transcriptional units within the P gene.(7) Of these, the N protein plays an important role in CDV detection, including methods based on RNA or protein, induction of neutralizing antibodies,(8) and in nucleocytoplasmic transport, nuclear localization/export.(9)
The nucleoprotein (N) and hemagglutinin (H) genes have been shown to induce specific humoral and cellular immune responses against CDV in mink.(10) These properties make the N protein a suitable target for the development of antiviral agents.
In a previous study, the monoclonal antibody (MAb) 1N8 was prepared by hybridoma technique using a truncated recombinant N protein (aa 277-471) of CDV as an immunogen. Further research revealed that the MAb 1N8 was capable of recognizing the native N protein of CDV with a high affinity.(11)
The single-chain fragment variable (scFv) is a small, engineered antibody in which the variable light chain (VL) and heavy chain (VH) of the antibody are connected by a short polypeptide linker. At present, the scFv molecule is considered a powerful tool for therapeutic antigen targeting in vivo.
In the current study, the scFv gene was obtained from the hybridoma cell 1N8 secreting the monoclonal antibody against N protein of CDV. After cloning, the framework regions and complementarity determining regions of the scFv gene were analyzed on the basis of the deduced amino acid sequences. The recombinant scFv protein was successfully expressed in Escherichia coli using the prokaryotic expression vector pET32a with a His-tag. Moreover, the binding activity and specificity of the scFv were determined by an indirect ELISA and competitive ELISA. Our aim is to reveal the basic sequence information of the MAb 1N8 and develop a practical procedure for the preparation of scFv to facilitate further viral detection research.
Materials and Methods
Strains and hybridoma cells
The hybridoma cell of the MAb 1N8 against CDV N protein was described in a previous study.(12) Escherichia coli Rosetta™ (DE3) plysS cells were obtained from Novagen (Madison, WI).
Primers for scFv gene
The primers for PCR amplification of the VL and VH genes of the MAb 1N8 were designed according to the degenerate primers described by Zhu and colleagues.(12)
The 5′ ends of the VH forward primers and VL reverse primers were modified to include KpnI and Xho I sites, respectively. In order to splice the genes VL and VH, a complementary overlapping sequence encoding a flexible linker of 12 amino acids (SSGGGGSGGGGS) was added to the 5′ ends of the VH reverse primers and VL forward primers, respectively. Detailed primer information is shown in Table 1.
S=G/C, R=G/A, K=G/T, M=A/C, Y=C/T, W=A/T, H=A/C/T, B=C/G/T, V=A/C/G, D=A/G/T, N=A/T/G/C.
RNA extraction, cDNA synthesis, and amplification
Total RNA was prepared from the hybridoma cells of the MAb 1N8 against N protein of CDV using the Ultrapure RNA Kit (CWBIO, Beijing, China) according to the manufacturer's instructions.
Using the extracted RNA as templates, the first strand cDNA synthesis was carried out with the SuperRT cDNA Kit (CWBIO), according to the manufacturer's instructions.
The VL and VH genes of the MAb 1N8 were respectively amplified by conventional PCR using synthesized cDNA as a common template. Briefly, the PCR reaction was performed in a 25 mL mixture containing 1 μL forward and reverse primer, 1 μL cDNA template, 0.25 μM each dNTP mixture, buffer, and 0.5 U ExTaq DNA polymerase. The amplification conditions comprised an initial denaturation at 95°C for 5 min, followed by 30 PCR cycles of 95°C for 30 s, 52°C for 30 s, 72°C for 30 s, and a final extension step of 72°C for 10 min. The amplified products of the VL and VH genes were purified using the Gel Extraction Kit (CWBIO).
Generation of scFv/1N8 gene
Using the purified VL and VH genes as templates, an overlap extension PCR (SOE-PCR) was performed for generation of the scFv gene of the MAb 1N8 (scFv/1N8). Briefly, the SOE-PCR reaction contained 100 ng of purified VL products and 100 ng of purified VH products, 0.25 mmol each dNTP mixture, 10× buffer, and 0.5 U ExTaq DNA polymerase. 35 PCR cycles were performed as follows: 95°C for 1 min, 50°C for 1 min, 72°C for 1 min, and a final extension step of 72°C for 10 min.
Prokaryotic expression of scFv/1N8 gene
The scFv/1N8 gene was digested using the restriction endonucleases KpnI and Xho I, and linked with the vector pET32a using T4 DNA Ligase at 16°C for 12 h. The ligation products were transformed into E. coli Rosetta™ (DE3) plysS cells, and the positive plasmids were selected to determine the sequence of the scFv/1N8 gene. Sequence analysis of the framework regions (FR) and complementarity determining regions (CDR) of the scFv/1N8 gene was carried out using the IMGT/V-QUEST program (v3.3.1) of the international immunogenetics information system(13) (available at
Purification of recombinant scFv/1N8 protein
The inclusion body of the recombinant scFv/1N8 protein was extracted from the lysate of the IPTG-induced host bacteria processed by supersonic waves. The extracted inclusion body proteins was purified by affinity chromatography using His Resin column (GE Healthcare, Piscataway, NJ), according to manufacturer's instructions. The purity and concentration of the protein were analyzed by SDS-PAGE. After electrophoresis, the gel was stained with the reagent rapid stain Reagent (CWBIO).
Refolding of denatured fusion protein
The purified scFv-His protein was dissolved in denaturing buffer and sonicated. Refolding was accomplished using a pH and urea gradient method(14) with the following modification. Briefly, purified scFv-His was dialyzed by increasing the pH (6 to 8.5) and decreasing the concentrations of urea (4 to 1 M) in the renaturing buffer containing 0.01 M PBS and 0.5 mM arginine at 4°C; buffer was changed every 12 h for 48 h. Oxidizing conditions were introduced by adding 0.3 mM oxidized glutathione (GSSG) and 3 mM glutathione (GSH) to the refolding buffer. The renatured scFv-His was finally dialyzed against 1 M urea containing 0.01 M PBS.
Antigen-binding activity of CDV-specific scFv
Indirect ELISA was performed using an ELISA assay to evaluate the antigen binding activity of purified scFv. Bound scFv was probed with His-tag and detected with TMB. The scFv was added as two-fold dilutions to pre-coated and blocked wells containing purified CDV NP protein incubated for 1 h at 37°C. The wells were washed three times. An anti-His-tag mouse monoclonal antibody (CWBIO) at 1:2000 was added to the wells (100 μL/well), and the plates were incubated at 37°C for 1 h. Plates were washed three times, an HRP-goat anti-mouse IgG (BoAoSeng Co., China) at 1:5000 was added to the wells, and then the plate was incubated at 37°C for 30 min. After three final washes, TMB (tetramethyl benzidine) was added to each well and the plates were incubated in the dark for 15 min at RT. The reaction was stopped by the addition of 1.25 M H2SO4, and the absorbance was measured at 450 nm using a microplate reader (Bio-Tek, Shoreline, WA). The experiment was performed thrice and each time ELISA measurements were done in triplicate.
Measuring activity of refolded scFv by competitive ELISA
The developed anti-CDV scFv was evaluated for CDV diagnosis in the format of competitive ELISA. The wells of ELISA plates were coated by the purified truncated CDV NP protein diluted in carbonate bicarbonate buffer (pH 9.4) in pre-determined dilution (0.05 mg/well). Wells were pre-blocked with 5.0% skimmed milk powder (Merck, Darmstadt, Germany). Antigen-coated plates were incubated with 50 μL of MAb and an equal volume (50 μL/well) of scFv per well with different concentration in PBST. The scFv-MAb mixtures were allowed to react with the antigen for 1 h at 37°C. The rest of the procedure was carried out exactly as described for the indirect ELISA above. The OD values were used to calculate the percent inhibition induced by scFv, using the formula percent inhibition=[1- (ODscFv/ODMAb)]×100, where ODscFv is the mean OD of wells with scFv, and ODMAb is the mean OD of wells with MAb alone.
Results
Amplification and analysis of scFv/1N8 gene
The VH and VL genes were amplified at about 360 and 400 bp by PCR using the cDNA from the hybridoma cells 1N8 as a template, and then about 750 bp of the scFv/1N8 gene was generated by SOE-PCR (Fig. 1). The nucleotide sequences and deduced amino acid sequences of three FRs and three CDRs of the H and L chains in the scFv/1N8 gene are shown in Table 2. Sequence analysis indicated that the VH gene of the scFv/1N8 gene showed 86% nucleotide identity with mouse immunoglobulin heavy chain variable region gene (Genbank accession no. NC_000078.6); the VL gene of the scFv/1N8 gene showed 96% nucleotide identity with mouse immunoglobulin kappa light chain variable region gene (Genbank accession no. AC_000028.1).
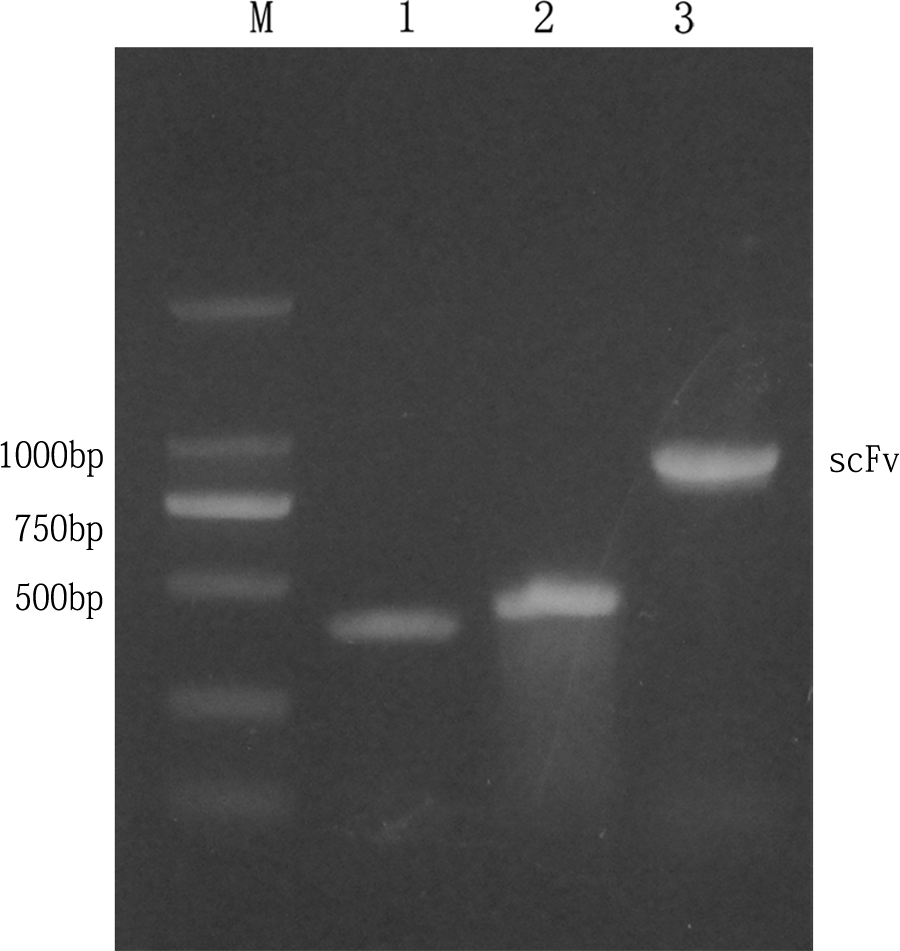
In vitro amplification of variable light and heavy chains of scFv/1N8. M, 2000 bp DNA ladder (Takara); lane 1, variable light chain (VL) of IgG of 400 bp; lane 2, variable heavy chain (VH) of IgG of 500 bp; lane 3, amplified products of scFv/1N8 gene by SOE-PCR (∼850 bp).
Expression and purification of scFv/1N8 gene
After cloning the scFv/1N8 gene into prokaryotic expression vector pET32a, the recombinant scFv/1N8 protein was successfully expressed at a high level in Escherichia coli; its molecular weight was approximately 43 kDa (Fig. 2). The high-purity recombinant scFv/1N8 protein was obtained by simple His Resin purification, according to the manufacturer's instructions (Fig. 2).

SDS-PAGE analysis of scFv fragment expression and purification in E. coli. Lane 1, un-transformed E. coli RosettaTM (DE3) plysS strains; lane 2, transformed, induced E. coli RosettaTM (DE3) plysS strains; lane M, prestained protein ladder (Fermentas); lanes 3–11, purified scFv by His Resin with different concentration iminazole; lane 8, 80 mM iminazole; lane 9, 120 mM iminazole; lane10, 160 mM iminazole.
Determination of scFv ability to bind to CDV NP antigen by indirect ELISA
The antigen binding activity of the purified scFv was measured by an indirect ELISA. Titration of scFv against the CDV NP protein revealed a concentration-dependent reduction of the optical density values, as shown in Figure 3A. No reactivity was observed with a lysate of E. coli, indicating the binding specificity of scFv.
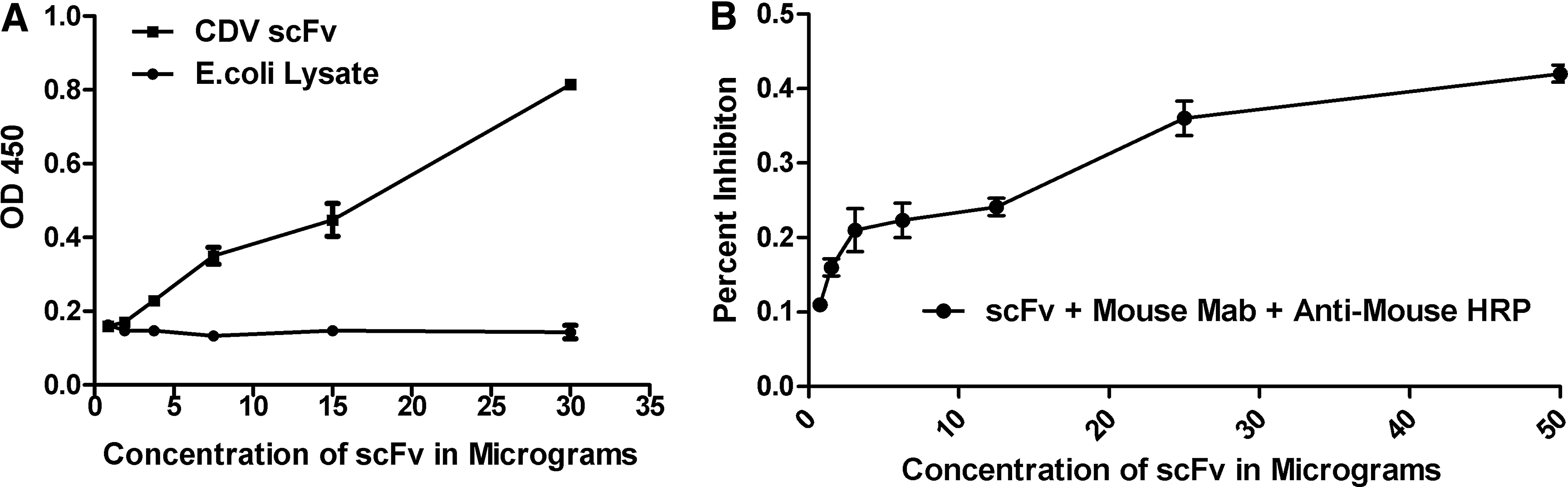
Analysis of antigen-binding affinity of scFv by indirect ELISA (
Competitive ELISA
Competition assay was performed to determine whether the scFv and mouse monoclonal antibody recognized the same epitope on CDV. The mouse MAb was incubated with varying 2-fold dilutions of scFv and detected using anti-mouse HRP. Detection with HRP-conjugated anti-mouse IgG showed an apparent decrease in the percent inhibition following the dilution of scFv with MAb 1N8, clearly indicating that both the scFv and MAb did compete for the same epitopes on the CDV N protein antigen (Fig. 3B).
Discussion
Canine distemper disease is an acute and highly contagious Canidae disease, which poses a serious threat to the fur animal industry because of its acute occurrence and high lethality. The emergence of new genotypes could contribute to the outbreak of CD.(15) At present, the main prevention and therapeutic measures of CD are vaccination and high immune serum.
The use of antibody is a powerful therapeutic tool against viral infections.(16) During the past decade, advances in recombinant antibody technology have led to the development of a large variety of engineered antibody molecules for research, diagnosis, and therapy with specificities out of reach of conventional antibody technology. Of these, scFv is the smallest unit of immunoglobulin molecule functioning in antigen-binding activities. In comparison to the parental antibody, scFv molecules have several advantages in clinical practices, including better penetration, lower immunogenicity, and therapeutic antigen targeting in vivo, as well as the ability to facilitate mass production in vitro. At present, numerous scFvs have been constructed against hapten, protein, carbohydrate, receptor, tumor antigen, and viruses. All these scFvs demonstrate good potential for use in several fields, including medical therapies and diagnostic applications.
In our study, the scFv gene of the MAb 1N8 against CDV N protein was cloned from hybridoma cells. Sequence analysis demonstrated that the VH and VL genes of the scFv/1N8 gene were composed of four framework regions and three complementarity determining regions; they showed high identity with the VH and VL (kappa) genes of mouse immunoglobulin published in GenBank. Many studies have proven that the scFv genes from different antibodies were successfully expressed in prokaryotic expression system using E. coli as host cells.(17) The method of indirect ELISA for detecting the binding activity of scFv has been established. The OD450 of scFv/1N8 could reach 0.8. However, OD450 of control protein is <0.2, which indicates high specific affinity.
In conclusion, we cloned the scFv gene from the hybridoma cells of MAb 1N8 against CDV N protein (aa 277-471) and expressed the recombinant scFv/6E6 protein in E. coli using the vector pET-32a with His-tag. Although the neutralization capacity of the scFv/1N8 antibody still needs to be tested in vivo/vitro, the scFv antibody scFv/1N8 has potential for producing a diagnostic or therapeutic reagent of CD disease. It provided a theoretic basis and opened a new avenue for the prevention and treatment of canine distemper disease virus infection.
Footnotes
Acknowledgments
The authors thank the members of our groups for helpful discussions, especially Dr. Zhang for advice and excellent technical assistance.
Author Disclosure Statement
The authors have no financial interests to disclose.