
Abstract
DAB2 interactive protein (DAB2IP), also known as ASK1-interacting protein-1 (AIP1), a novel member of the RasGTPase-activating protein family, plays a key role in tumor suppression during cancer progression and is highly expressed in vascular smooth muscle cells (VSMCs) and endothelial cells (ECs). To further explore its function as a cancer suppressor, in this study, we immunized BALB/c mice with synthesized human DAB2IP polypeptide and obtained a novel monoclonal antibody (MAb) against human DAB2IP. A stable strain of hybridoma was screened and successfully established by the hybridoma technique. The immunohistochemistry, immunocytochemistry, and Western blot analysis revealed that the MAb was directed against human DAB2IP with high specificity. Therefore, this MAb may be a useful tool and facilitate studies on tumorigenesis associated with DAB2IP.
Introduction
T
In summary, the above studies indicated that DAB2IP might be a suppressor of tumorigenesis, even a biomarker of tumors. In order to further elucidate the function of human DAB2IP in tumorigenesis, in this study, we used a synthetic peptide, the disabled homolog 2-interacting protein isoform 3, as immunogen and generated a monoclonal antibody (MAb) against human DAB2IP by the hybridoma technique. Using this MAb, we successfully detected DAB2IP expression in many cancer cell lines and tissues and normal cell lines and tissues by Western blot, immunocytochemistry, and immunohistochemistry.
Materials and Methods
Mouse preparation
Six- to 8-week-old, purebred female BALB/c mice were obtained from the Medical Animal Center of Sichuan University (Chengdu, China). All animal procedures were approved by the Sichuan University Committee on Animal Research.
Immunogen preparation
DAB2IP polypeptide (CRTRSAERPALPPQRSHL) was synthesized by (ChinaPeptides Co., Shanghai, China) as immunogen. Two mg lyophilized BSA were dissolved in 200 mL conjugation buffer (0.1 M 2-(N-morpholino) ethane sulfuric acid (MES). Two mg synthesized hDAB2IP polypeptide were then added in 500 mL conjugation buffer as a carrier-peptide solution. 100 μL EDU solution with a concentration of 10 mg/mL (diluted by DDH2O) was immediately added to the carrier-peptide solution. After incubating at room temperature for 2 h, the conjugate was purified by a desalting column.
Immunization of mice and cell fusion
The female BALB/c mouse (6–8 weeks, about 20 g) was subcutaneously immunized with 100 μg human DAB2IP polypeptide emulsified with Freund's complete adjuvant (CFA, Sigma, St. Louis, MO). To enhance immunity, subcutaneous injection with the same dose and method was repeated three times within 2 mo. Before the last injection, blood was drawn from the mouse tail vein and the serum was isolated. The serum titers were analyzed by enzyme-linked immunosorbant assay (ELISA). The results showed that the mouse serum was highly immunoreactive to the immunogen. After the final booster injection, the mouse was killed by cervical dislocation, and spleen cells were collected for fusion with SP2/0 cells using polyethylene glycol (PEG) 4000 at a splenocyte-myeloma cell ratio of 10:1. The fused cells (hybridomas) were put into 96-well cell culture plates, which contained the normal BALB/c mouse feeder cells and HAT (Sigma) dissolved into RPMI-1640 medium.
Hybridoma cell screening
To obtain positive clones producing anti-DAB2IP antibody, ELISA was used for screening and identification. Two 96-well microtiter plates were prepared and coated by synthesized human DAB2IP polypeptide as coating antigen with 1 μg/mL. The plates were blocked by 1% casein 120 μL in each well at 4°C overnight. 50 μL/well of hybridoma supernatant were added into the plates as primary antibody whereas RPMI-1640 medium was used as negative control and incubated at 37°C for 1 h. After washing three times with PBST, 100 μL/well of goat anti-mouse IgG conjugated with HRP were added into the plates as secondary antibody (ZSGB-BIO) and incubated at 37°C for 1 h. The plates were washed with PBST and 100 μL tetramethyl-benzidine (TMB) substrate was added at 37°C within 15 min. Two M H2SO4 was used for stopping the reaction. The optical density absorbance value (OD) was detected by ELISA reader (Bio-Rad, Hercules, CA) at 450nm. The positive clones were selected and subcloned into 96-well plates, and the supernatants of positive clones were detected by ELISA. The human DAB2IP antibodies were determined in the same way.
Generation and purification of ascites
Five 10-week-old BALB/c mice were prepared and intraperitoneally injected with liquid paraffin (500 μL/mouse). Then, each mouse was injected peritoneally with 5×106 hybridoma cells, producing the anti-DAB2IP antibodies after 1 week. After 10–14 days, the mice were killed by cervical dislocation. Then ascites were collected and stored at 4°C with 1% sodium azide. The ascites were purified by ammonium sulfate precipitation, following further depuration with protein-G affinity chromatography (Pharmacia Biotech, Uppsala, Sweden).
Detection of ascites titer by ELISA
In this step, the method was similar to that described in the hybridoma cell screening section (see above).
Western blot analysis
The lysates of human PC-9, H460, A549, EC-109, TE-1, H1299 cancer cells and normal cells HUVEC, MRC-5, HBE were separated by 10% SDS-PAGE. The samples were electro-transferred onto a polyvinylidene fluoride (PVDF) membrane and blocked by 5% skim milk in TBST at room temperature for 1 h. The ascites (anti-DAB2IP) with 1:500 dilutions were added onto the membrane at 4°C for 48 h. After 48 h and washing the membrane three times with TBST, the horseradish peroxidase-conjugated secondary antibody was used at a dilution of 1:5000. The protein of interest was detected by chemiluminescence.
Immunocytochemistry and immunohistochemistry
A375, B16, and Hela cells were seeded onto different cover slips, which were included in the 6-well plates. After cell adhesion, the seeded cells were fixed with 4% paraformaldehyde (PFA) at 20°C for 15 min. Then PFA was removed from the plates and the slips were washed three times with PBST. 0.1% Triton X-100 was used for cell permeation for 10 min. Endogenous peroxidase activity was inactivated by 3% H2O2 for 15 min. 0.25% pancreatin was used for antigen retrieval at 37°C for 15 min. All cells were blocked by solution A (goat serum) for 15 min. Then the ascites (20 mg/mL) of anti-human DAB2IP MAb as primary antibody was added and incubated at 40°C overnight, using PBST as negative control. After being washed with PBST, a secondary antibody, solution B (anti-mouse IgG antibody) was added and incubated at room temperature for 15 min, then washed again. Solution C (HRP-coupled streptavidin) was used for conjunction to solution B. At the end of the last washing, diaminobenzidine (DAB) chromogen system was used as a detector to recognize solution C, and the reaction was completed within 5 min. The slips were counterstained with Mayer's hematoxylin, dehydrated with the concentrations of ethanol from 70–100%, transparentized with dimethylbenzene, and finally mounted. For IHC staining, paraffin-embedded specimens of pulmonary adenocarcinoma, cervical cancer, bone tumor, and spleen were prepared. Paraffin sections were put into an antigen retrieval solution at 95°C for 30 min and then dewaxed in xylene and rehydrated with distilled water. The endogenous peroxidase activity and non-specially antigen of sections was respectively blocked by 3% H2O2 and goat serum. The primary antibody of DAB2IP was added onto slides with 1:100 and incubated overnight at 4°C. The following reactions were performed by ABC systems (ZSGB-BIO) and DAB stain (ZSGB-BIO), counterstained by hematoxylin. All of these results were analyzed with a light microscope.
Affinity constant measurement
The affinity constant of the anti-DAB2IP MAb was measured by non-competitive enzyme immunoassay. The 96-well plate was coated with three different concentrations of synthesized human DAB2IP polypeptide (2.0, 1.0, 0.5 mg/L); doubling diluted anti-DAB2IP antibody was added and incubated at 37°C for 1 h, followed by washing. Then the plate was incubated with secondary antibody (anti-mouse IgG antibody) for 1 h at 37°C, and TMB (tetramethylbenzidine) was added for color development. The optical density (OD) values were detected at 450nm. The curves were made with OD value as Y axis, whereas the concentration logarithm of anti-DAB2IP MAb was X axis (Fig. 1). The upper platform stage of curves was designed at OD100. Followed find out the OD50 value versus logarithm of MAb concentrationand turn the logarithm value into concentration, (Ab)t1 (Ab)t2 (Ab)t3, respectively. The affinity constant of antigen-antibody interaction was designed by the equation Kaff=(n–1)/2(n(Ab′)t – (Ab)t); when the concentration ratio of the antigen was 1:2, K1=1/2(2(Ab)t1 – (Ab)t2), K2=1/2(2(Ab)t2 – (Ab)t3), and K3=3/2(4(Ab)t1 – (Ab)t3) when the ratio was 1:4. The final K value was the average of the three results, which was our anticipated result.
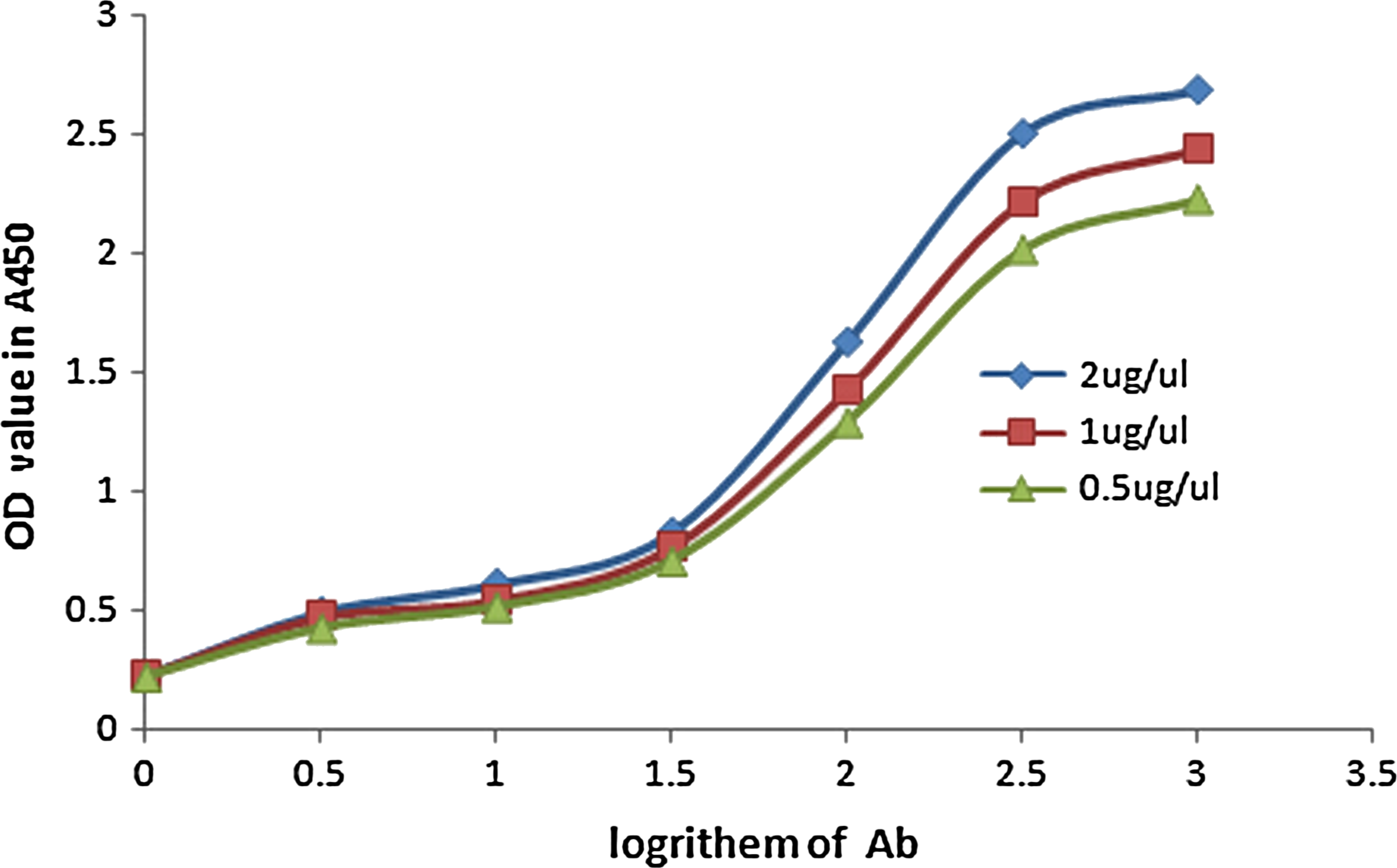
Affinity curves of anti-DAB2IP MAbs.
Cell line and culture conditions
All human lung cancer cells and normal cells were cultured in media glucose Dulbecco's modified Eagle's medium (HyClone, Shanghai, China) and A375, B16, HeLa cells in RPMI-1640 supplemented with 10% fetal bovine serum, 100 U/mL penicillin, and 100 mg/mL streptomycin at 37°C in a humidified atmosphere containing 5% CO2.
Results
Screening of anti-DAB2IP MAb-producing hybridoma positive clone
Five BALB/c mice were immunized with human DAB2IP protein and anti-DAB2IP MAb was obtained by hybridoma technology. The hybridoma cell clones producing anti-DAB2IP MAb were selected by ELISA using synthesized human DAB2IP polypeptide as antigens and RPMI-1640 medium as negative control. The stable cell line that could stably produce anti-DAB2IP MAb was established.
Generation of large-scale MAb
The stable cell line producing anti-DAB2IP MAb was injected into the abdomen of BALB/c mice. Then ascites were obtained after 10 days. The isotype of heavy chain and light chain of the mouse anti-DAB2IP MAb were IgG2a and k, respectively.
Western blot analysis
The specificity of anti-DAB2IP MAb to human DAB2IP protein was detected with Western blot (Fig. 2). Each lane was the whole protein lysate of human lung cancer, esophagus cancer, and normal cells, respectively. The molecular weight of human cell lysate band was more than 110 kDa (Fig. 2B), due to the heavy chain modification with O-linked carbohydrates of anti-DAB2IP MAb. The result indicated that the anti-DAB2IP MAb was highly specific against the human DAB2IP protein. The human DAB2IP detected different expressions in different cell lines, with a high level in A549 and little expression in HUVEC, H460, and H1299 (Fig. 2).

Western blot results of DAB2IP protein expression with anti-DAB2IP MAb. While not positive in HUVEC, the DAB2IP protein expression was up-regulated in A549 compared to normal (
Immunocytochemistry
Positive signals were detected in the cytoplasms of A375, Hela, and B16 cells as well as (skeletal) tissues by immunocytochemistry. These results confirmed that DAB2IP proteins were expressed in these cells and tissues. In contrast, no positive expressions were observed in PBS groups as controls (Fig. 3).

Immunohistochemistry analysis of paraffin-embedded cervical cancer (a), bone tumor (b), pulmonary adenocarcinoma (c), spleen (d) (
Immunohistochemistry
The high positive signal results were detected respectively in cervical cancer, bone tumor, pulmonary adenocarcinoma, and spleen (Fig. 3B).
Affinity constant of anti-DAB2IP MAb
The affinity constant of anti-DAB2IP MAb was 2.8×108 L/mol by non-competitive enzyme immunoassay, as shown in Figure 1.
Discussion
DAB2 interactive protein (DAB2IP), a novel Ras GTPase-activating protein family member, is expressed in human brain and the soma, and is closely related to the developing cerebral cortex, regulating neuronal migration.(1,2,4,5) Recent studies on DAB2IP were mainly focused on cancer suppression. Some studies showed DAB2IP protein was down-regulated in prostate cancer (PCa) and lung cancer, breast cancer, and hepatocellular carcinoma. DAB2IP gene aberrant methylation was detected in breast cancer, PCa, lung cancer, and gastrointestinal tumors.(6–10) Loss of DAB2IP expression caused cancer cell metastasis by controlling a step of epithelial to mesenchymal transition (EMT), the human PCa grew rapidly in vivo, and the development of multiple lymph nodes and distant organ metastases.(6,9,11) DAB2IP could regulate various cell survival and apoptosis signal pathways such as PI3K-Akt, wnt and NF-κB, ASK1-JNK.(6,13) DAB2IP also could inhibit RAS pathway by its Ras GTPase activity.(14) Knockdown of DAB2IP led to increased cell proliferation and S-phase cell distribution, activation of ERK and Akt, enhanced cell migration, and invasion ability in urothelial carcinoma cell lines of the bladder. Five-year survival rates significantly improved with high expression of DAB2IP in urothelial carcinoma of the bladder after surgery.(15) Previous studies showed that DAB2IP expression was distinctly lower in pancreatic cancer tissues, and wild-type KRAS gene of pancreatic cancer patients than in adjacent tissues and normal pancreatic tissues.(16) In prostate cancer, DAB2IP knockdown (KD) cells can result in chemoresistance by regulating the Egr-1/Clusterin signaling network.(19) Some research results indicate that human DAB2IP is down-regulated in hepatocellular carcinoma, and we considered the possibility that DAB2IP could be a cancer suppressor.(18–20) The fact that DAB2IP correlated with cancer stem cell properties (CSC) was observed in some studies.(21) AR (androgen receptor) was active abnormally in the DAB2IP(-/-) mice model of prostate and led to hyperplastic epithelia. DAB2IP can curb cancer cell survival by interfering with AR nuclear translocation or phosphorylation.(22) The above results gave us a clue that DAB2IP might be a suppressor or biomarker in tumorigenesis.
In our research, to explore the expressions and functions of human DAB2IP (hDAB2IP) in tumorigenesis, a novel anti-hDAB2IP MAb was successfully established through the hybridoma technique. Western blot, immunocytochemistry, and immunohistochemistry results showed that anti-hDAB2IP MAb could recognize the natural hDAB2IP protein specifically. Using this MAb, we have detected hDAB2IP expressions in diverse human cancer cells and normal cell lines and tissues by Western blot, immunocytochemistry, and immunohistochemistry assays. Our results indicated that obvious expressions of the hDAB2IP protein were observed in many different cancer cell lines, such as A375, B16, Hela cells, whereas there were faint or absent expressions in H460, H1299 cancer cells. The expressions of hDAB2IP protein were observed in normal cells MRC-5 and HBE but not in HUVEC. Meanwhile IHC results indicated positive expressions in many tissues such as cervical cancer, bone tumor, pulmonary adenocarcinoma, and spleen. Therefore, our results showed there was no significant difference in hDAB2IP protein expression between cancer cells/tissues and normal cells/tissues. It was not coincidental that some researchers reported that the DAB2IP protein might be a cancer suppressor.(6,11,13) Our results indicated that DAB2IP protein were highly expressed in A549 and PC-9 cancer cell lines and pulmonary adenocarcinoma tissues, which indicated that hDAB2IP might be a biomarker of pulmonary adenocarcinoma.
According to previous results and our results, DAB2IP might be a factor associated with tumorigenesis and a potential biomarker or therapeutic target. Additional studies should be carried out for further mechanism exploration using the anti-DAB2IP MAb as a new tool.
Footnotes
Acknowledgments
This research was supported by the members of the Department of Immunology, West China School of Preclinical and Forensic Medicine (Sichuan University, Chengdu, China). The human lung cancer and normal cell lines were provided by the Department of Thoracic Oncology, West China School Hospital of Sichuan University. The tissue was supplied by the Department of Immunology, West China School of Preclinical and Forensic Medicine (Sichuan University, Chengdu, China). We thank Dr. Nan Li for review of the manuscript.
Author Disclosure Statement
The authors have no financial interests to disclose.