
Abstract
IgE mainly activates cells via two receptors, FcɛRI and FcɛRII. Blocking antibodies against and animals genetically targeted for these receptors have been successfully used to distinguish between these two activating pathways. In the present study, we investigated whether our newly established anti-ovalbumin (OVA) monoclonal IgE OE-2 induced FcɛRII-dependent activation, but not FcɛRI-dependent activation in vivo and in vitro, in contrast to the previously established anti-OVA IgE OE-1, which stimulated FcɛRI and FcɛRII. The FcɛRI-mediated degranulation of RBL2H3 cells and passive systemic anaphylaxis in mice were induced by OE-1 but not OE-2. On the other hand, the production of nitric oxide by rat peritoneal macrophages and the primary antibody response in mice against co-injected OVA, which were mediated through FcɛRII, were induced and enhanced by OE-1 and OE-2. Differences in the epitopes recognized by OE-1 and OE-2 may partially explain why OE-1, but not OE-2, triggered FcɛRI-dependent activation. OE-1 bridged FcɛRI through effective aggregation with OVA, whereas OE-2 crosslinked the receptor strongly and only moderately upon the addition of an anti-kappa antibody and polymerized OVA, namely, an OVA-conjugated resin, respectively, resulting in degranulation. Our results offer a novel approach for determining the relative importance of FcɛRI and FcɛRII in various IgE-dependent responses by using OE-1 and OE-2.
Introduction
T
IgE is known to modify immune responses through other IgE receptors, including the low affinity IgE receptor FcɛRII (CD23),(5) FcγRIV,(6,7) and galectin-3.(8) FcɛRII belongs to the lectin family and is ubiquitously expressed on various cells.(5) The immune complex comprising IgE and the recognized antigens binds to the receptor and induces the transduction of an activation signal from Fyn.(9) Macrophages have been shown to produce nitric oxide (NO) in vitro in a manner that is dependent on FcɛRII when stimulated with IgE and the recognized antigens,(10) which has been associated with parasite rejection.(11) IgE-dependent immune responses against the recognized antigens were previously reported to be mediated in vivo through FcɛRII expressed on B cells(12) and have been associated with the role of the receptor in controlling serum IgE levels.(13) These findings indicate that FcɛRII does not directly mediate acute allergic reactions, but exacerbates allergic responses indirectly by enhancing sensitization against the recognizing antigens.
FcɛRI-(14) and FcɛRII-(12) knockout mice and antibodies against these receptors(15,16) have been used successfully to determine the exclusive contribution of each of these receptors to various IgE-dependent phenomena, including allergic responses. Experiments that use knockout mice and antibodies in parallel are recommended in order to increase the reliability of the findings obtained. Research quality may be compromised when either or both of these two methods cannot be used. Therefore, additional methods need to be developed in order to determine whether the effects of IgE are FcɛRI- and FcɛRII-dependent. For example, a monoclonal IgE capable of inducing only FcɛRI- or FcɛRII-mediated signaling may be advantageous for determining the roles of each receptor in IgE-mediated responses against the recognized antigens.
Therefore, we established a new hybridoma producing anti-OVA monoclonal IgE OE-2. OE-2 with OVA induced FcɛRII-mediated responses, but not FcɛRI-mediated responses. The characteristics of OE-2 in comparison with the previously produced anti-OVA IgE OE-1,(17) which induced FcɛRI- and FcɛRII-mediated responses, were evaluated, and the possible applications of these two antibodies in future research are discussed.
Materials and Methods
Hybridomas, antibodies, and cell lines
Hybridomas producing mouse anti-OVA monoclonal IgE (OE-2) and anti-OVA monoclonal IgG1 (O1-8) were newly established by the classical method using PEG1500 (Roche Diagnostics, Mannheim, Germany) following the manufacturer's instructions.(17,18) Mouse anti-OVA monoclonal antibodies including OE-1, OE-3, O1-10, OA-4, and those described above, were purified with affinity chromatography followed by ultrafiltration, as reported previously.(18) The following antibodies were also purchased: alkaline-phosphatase-conjugated anti-IgE and anti-kappa light chain antibody (Southern Biotech, Birmingham, AL); alkaline-phosphatase-conjugated anti-biotin (Vector Laboratories, Burlingame, CA); alkaline-phosphatase-conjugated anti-mouse IgG1 (X56) and IgG2a (R19-15) (BD Bioscience, San Jose, CA); and standard mouse IgE SPE-7 (Sigma-Aldrich Fine Chemicals, St. Louis, MO). Rat basophilic leukemia RBL2H3 cells were obtained from the Cell Resource Center for the Biomedical Research Institute of Development, Aging, and Cancer, Tohoku University, Sendai, Japan.
Animals
Male DBA/1J mice (8–12 weeks old; Charles River Laboratories, Yokohama, Japan) were housed and maintained in a controlled environment and given standard chow and water. All animal experiments were approved by the Animal Ethics Committee of Kobe Pharmaceutical University.
Enzyme-linked immunosorbent assays to identify differences in affinities for OVA and epitopes recognized between OE-1 and OE-2
ELISAs to determine the affinities of OE-1 and OE-2 for OVA and differences in the recognized epitopes have been described previously.(19) In brief, OVA-coated ELISA plates were incubated with the indicated concentrations of OE-1 or OE-2. In order to evaluate their affinities, the plates were incubated with alkaline phosphatase-conjugated anti-mouse IgE. Alternatively, the plates were further incubated with biotinylated OE-1 or OE-2, followed by incubation with alkaline phosphatase-conjugated anti-biotin antibodies in order to determine the recognized epitopes. In both cases, alkaline phosphatase activities in the plates were colorimetrically detected using p-nitrophenyl phosphate (Sigma-Aldrich).
Measurement of binding ability of OE-1 and OE-2 to recombinant FcɛRI and FcɛRII in ELISA
The wells of ELISA plates were coated with 0.5 μg/100 μL phosphate-buffered saline (PBS) of recombinant mouse FcɛRI (Fc Tag) (Sino Biological, Beijing, China) or recombinant mouse CD23 (FcɛRII, R&D Systems, Minneapolis, MN) at 4°C overnight. The wells were washed with 0.5% Tween-20-containing PBS and blocked with Superblock Blocking buffer in PBS (Thermo Scientific, Rockford, IL). After washing, the indicated concentrations of OE-1, OE-2, and O1-10 were added to the wells and incubated for 1 h at room temperature. The IgE or IgG1 antibodies that bound to the receptors were then detected colorimetrically using p-nitrophenyl phosphate following incubation with the alkaline phosphatase-labeled anti-kappa light chain antibody.
Degranulation of anti-OVA IgE-sensitized RBL2H3 cells with OVA, anti-kappa light chain antibody, and OVA-conjugated agarose
RBL2H3 cells (0.25 × 106/mL) in RPMI1640 medium (Wako Pure Chemical Industries, Osaka, Japan) containing 10% fetal bovine serum (Biowest SAS, Nuaillé, France), penicillin (100 U/mL)-streptomycin (100 μg/mL) (Wako Pure Chemical), and the indicated concentrations of OE-1 and OE-2 were cultured for 24 h. These sensitized cells were washed with PIPES buffer (25 mM PIPES [pH 7.2], 119 mM NaCl, 5 mM KCl, 5.6 mM glucose, 0.4 mM MgCl2, 1 mM CaCl2, and 0.1% (w/v) bovine serum albumin). The cells were then stimulated for 20 min with buffer containing 5 μg/mL of OVA, the indicated concentrations of the anti-kappa light chain antibody, and 5% or 25% v/v OVA-agarose, which was produced with OVA and cyanogen bromide-activated-Sepharose 4B (Sigma-Aldrich). The degranulation ratios of β-hexosaminidase secreted into the supernatants were determined by a colorimetric reaction using p-nitrophenyl-N-acetyl-β-D-glucosaminide (Sigma-Aldrich).
Passive systemic anaphylaxis and de novo anti-OVA antibody production induced with immune complex injection
Passive systemic anaphylaxis was induced by an intravenous injection of OE-1 or OE-2 (200 μg), followed by an intravenous injection of OVA (50 μg) the next day. The indicator of anaphylaxis, namely, a decrease in rectal temperature, was monitored for 1 h from the antigen injection. In order to measure the production of de novo anti-OVA and anti-dinitrophenyl-keyhole limpet hemocyanin (DNP-KLH, 200 μg; LSL Co., Tokyo, Japan) antibodies, OE-1, or OE-2 (200 μg) was mixed with OVA (50 μg) and DNP-KLH (200 μg) and then injected. Sera were collected on day 11 to measure de novo anti-OVA and anti-DNP-KLH antibody production as indicators of immune responses.
Measurement of anti-OVA and anti-DNP-KLH IgGs in sera
Anti-OVA IgG, IgG2a, and IgG1 levels and anti-DNP-KLH IgG, IgG2a, and IgG1 levels in sera collected on day 11 were determined by ELISA using OVA- and DNP-KLH-coated plates, as described previously.(18)
Determination of NO production from rat peritoneal macrophages
Rat peritoneal macrophages were obtained as described previously.(20) These cells were stimulated with the indicated concentrations of OE-1, OE-2, and OVA for 20 h in RPMI1640 medium containing fetal bovine serum and antibiotics. NO concentrations in the cultured supernatants were determined using Griess reagent,(21) as described previously.(22)
Statistical analysis
The Mann–Whitney U test was used to compare differences between two independent groups. A one-way ANOVA with Tukey's post-hoc test was used for multiple comparisons. A p value of less than 0.05 was considered significant.
Results and Discussion
Differences in epitopes recognized by OE-1 and OE-2, which had similar affinities for OVA
SDS-PAGE followed by Coomassie brilliant blue staining and Western blot analysis with anti-IgE revealed that OE-1 and OE-2 showed similar mobilities on SDS-PAGE and reacted with anti-IgE (Fig. 1A). The binding of OE-1 and OE-2 to OVA at concentrations of 0.01–10 μg/mL was similar in ELISA using an anti-IgE antibody as a detection antibody (Fig. 1B). The binding of biotinylated OE-1 to coated OVA was markedly reduced by the pretreatment of the plate with OE-1, while it was weakly inhibited by the pretreatment with OE-2 (Fig. 1C). The binding of biotinylated OE-2 to coated OVA was markedly decreased by the OE-2 pretreatment, but only moderately by the OE-1 pretreatment (Fig. 1C). These results suggest that OE-1 and OE-2 bound to different or partially overlapping, but not the same, epitopes on OVA.

OE-1, but not OE-2, induced FcɛRI-mediated reactions in vivo and in vitro, with variations being attributed to differences in their recognized epitopes. (
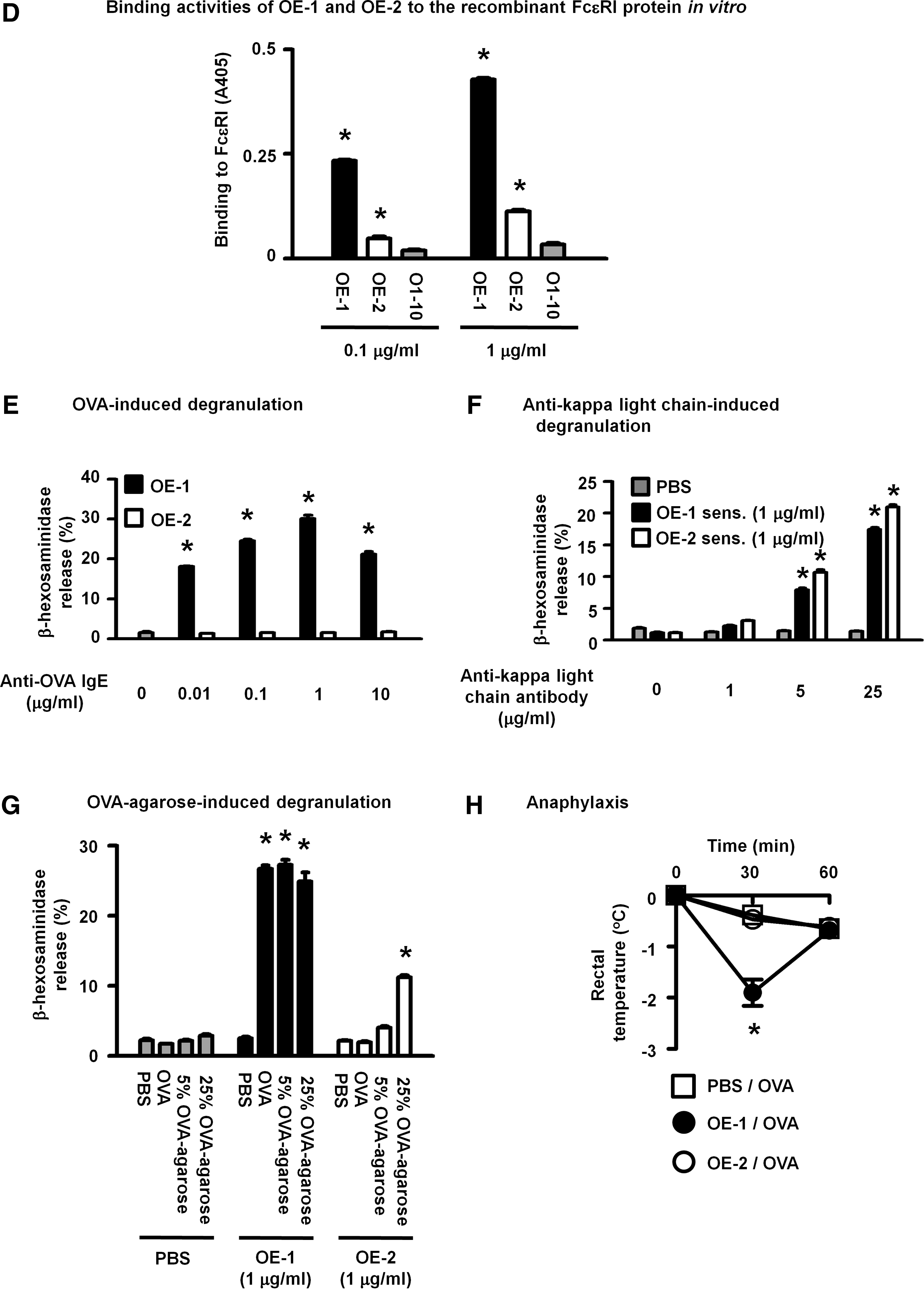
OE-1, but not OE-2, induced FcɛRI-dependent responses, namely, OVA-dependent degranulation of RBL2H3 cells and anaphylactic reactions in mice
OE-1 and OE-2 bound more strongly to recombinant FcɛRI than O1-10, an anti-OVA IgG1 monoclonal antibody, in ELISA (Fig. 1D). OE-1 had a higher affinity to the receptor than OE-2. The sensitization of RBL2H3 cells with OE-1 at concentrations greater than 0.01 μg/mL enabled cells to degranulate in response to OVA stimulation (Fig. 1E). On the other hand, OVA did not induce the degranulation of cells sensitized with OE-2 at any of the concentrations tested (Fig. 1E). In contrast to OVA, the anti-kappa light chain polyclonal antibody induced degranulation to the same extent in OE-1- and OE-2-sensitized RBL2H3 cells (Fig. 1F), indicating that OE-2 bound to FcɛRI and induced a stimulatory signal in cells through the aggregation of receptors following the addition of the anti-kappa light chain antibody. β-hexosaminidase was significantly released from OE-1- and OE-2-sensitized RBL2H3 cells upon stimulation with OVA-resin that mimicked a multivalent antigen (Fig. 1G). In spite of antigen valence, immune complexes with OE-2 and OVA may cause insufficient FcɛRI aggregation and, thus, did not induce a strong degranulation response. This result suggests the aggregation rate of FcɛRI by anti-OVA IgE was dependent on the recognized epitopes of OVA. The induction of an FcɛRI-dependent response by OE-1, but not OE-2, as shown in the in vitro experiments, was also observed in vivo, namely, anaphylaxis in OE-1-sensitized, but not OE-2-sensitized, mice (Fig. 1H). In contrast to OE-1, OE-2 lacked the ability to induce an FcɛRI-mediated activation signal in mast cells with OVA stimulation in spite of its ability to bind to the receptor.
OE-1 and OE-2 both induced FcɛRII-dependent responses, namely, OVA-dependent NO production by rat peritoneal macrophages and enhanced de novo anti-OVA IgG production in mice
Similar to FcɛRI, OE-1 strongly and OE-2 moderately bound to recombinant FcɛRII in ELISA (Fig. 2A). OE-1 and OE-2 both induced the production of NO by macrophages with OVA in a concentration-dependent manner (Fig. 2B) through a CD23-dependent mechanism.(10) The in vivo CD23-dependent enhancement in sensitization,(12) which was observed as increases in de novo anti-OVA IgG and Th2-dependent anti-OVA IgG1 production with concomitant decreases in Th1-dependent anti-OVA IgG2a production, was induced by injections of either OE-1 or OE-2 complexed with OVA, while the enhancing effect of OE-2 was slightly weaker than that of OE-1 (Fig. 2C). Neither antibody affected the primary antibody response against the non-related antigen DNP-KLH, as evidenced by the similar levels of anti-DNP-KLH IgG, IgG2a, and IgG1 in mice injected with OVA/DNP-KLH, OVA/DNP-KLH with OE-1, and OVA/DNP-KLH with OE-2 (Fig. 2C).
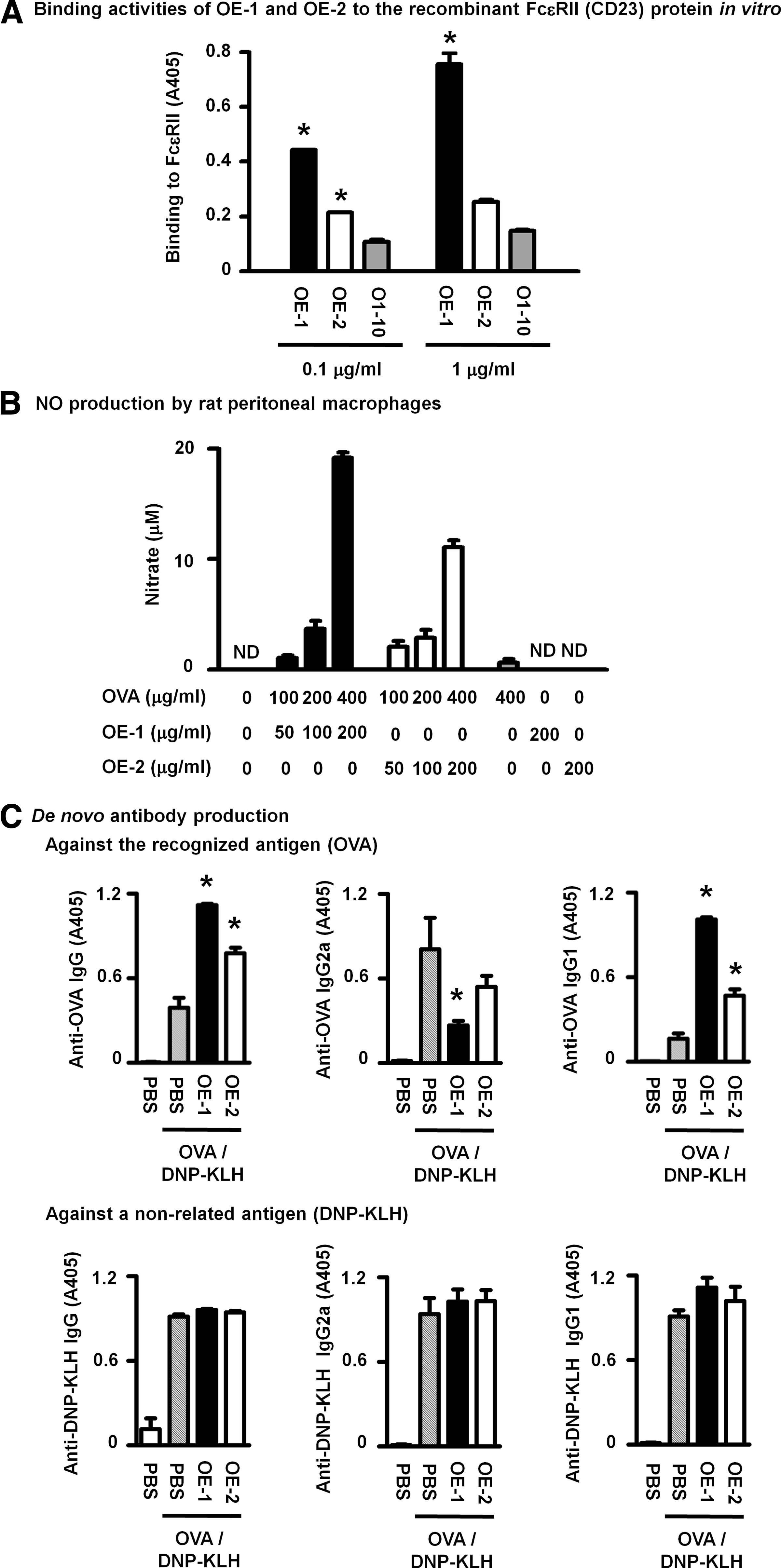
OE-1 and OE-2 similarly induced FcɛRII-mediated reactions in vivo and in vitro. (
Of the many monoclonal IgEs, two commercially available antibodies, anti-DNP IgE SPE-7(23) and anti-OVA IgE 2C6,(7) have been commonly used in research to elucidate the functions of IgE. SPE-7 has been shown to strongly induce an activation signal from FcɛRI(24) and FcɛRII(25) receptors with the multivalent antigens, DNP-conjugated proteins. The FcɛRI-dependent reaction induced by 2C6(26) appears to be weaker than that by SPE-7, and has been attributed to the low valence of the antigen OVA. To the best of our knowledge, it currently remains unknown whether 2C6 induces an activation signal through FcɛRII. OE-2 has the unique ability to induce FcɛRII-mediated responses without inducing FcɛRI-mediated responses, and, thus, is useful for evaluating the role of FcɛRII, especially in in vivo responses with an IgE/antigen stimulation, with the advantage of excluding the possible contribution of FcɛRI. In other words, in order to examine the role of FcɛRII, wild-type mice may be substituted for FcɛRI-knockout mice when OE-2 is used. For example, a previous study reported that FcɛRI did not participate in the enhancement of antigen-specific immune responses by IgE through FcɛRII in mice genetically lacking the expression of FcɛRI as well as FcγRI and FcγRIII.(12) We herein demonstrated more directly that the IgE-mediated enhancement of antigen-specific immune responses was inducible in the absence of FcɛRI-mediated activation by using OE-2, which is consistent with previous findings. More detailed information on IgE-dependent adjuvant effects will be obtained under these experimental conditions using various pharmacological inhibitors and signaling molecule-deficient mice. In addition, in vivo analyses may be easier to conduct using OE-2, due to its receptor selectivity, in order to clarify the mechanisms underlying IgE-CD23-dependent phenomena, including macrophage activation(10) and IgE/antigen transcytosis of epithelial cells,(27,28) proposed only based on in vitro findings. Anti-IgE 8D6, which binds to FcɛRII-associated IgE, but not to FcɛRI-associated IgE, was recently produced.(29) A comparison of the activation mechanisms of FcɛRII by OE-2/OVA and 8D6/IgE may be important for understanding the molecular characteristics of FcɛRII associated with the induction of subsequent intracellular signaling and reactions.
The lower affinity of OE-2 appeared to be responsible for its inability to induce FcɛRI-mediated responses upon OVA stimulation. However, the FcɛRI-mediated massive degranulation of RBL2H3 cells sensitized with OE-2 using the anti-kappa antibody demonstrated the sufficient binding of OE-2 to FcɛRI in the cell culture and, in turn, discouraged this idea. Furthermore, the affinity of OE-2 to FcɛRII was lower than that of OE-1 in spite of the strong activation of the receptor by the antibody and OVA. These results indicate that the binding of OE-2 to the receptors was moderate, but sufficient to induce receptor-mediated responses upon adequate stimuli such as the anti-kappa antibody for FcɛRI. Therefore, the different epitopes recognized by OE-1 and OE-2 may determine the activation abilities of these antibodies through FcɛRI. Immune complexes have many of the Fc portions of these antibodies. The distances of these portions are considered to be associated with receptor activation and differ among antibodies and antigens. The characteristics of antigens, including their sizes, recognized epitopes, and valencies, may be the factors influencing these distances. On the other hand, experiments using immune complexes composed of antibodies and natural antigens or the same antigen inserted into various spacers clarified that the favorable distance of binding epitopes of IgE for the crosslinking of FcɛRI was rigid and between 5 and 10 nm.(30) Although direct information on the suitable distance for FcɛRII is not currently available, the binding manners of IgE to FcɛRI and FcɛRII differ.(31) Moreover, conformational plasticity at the IgE-binding site of FcɛRII was previously reported to be extreme.(32) Therefore, one possibility is that OE-1 immune complexes had Fc portions that generated suitable distances for the stimulation of both receptors in response to OVA (containing oligomeric OVA). In contrast, OE-2 immune complexes had portions that generated acceptable distances for the FcɛRII stimulation, but not for FcɛRI. The induction of degranulation of OE-2-sensitized RBL2H3 cells by OVA-resin may also support this idea. The association between OE-2 and soluble OVA (oligomeric OVA) did not form an immune complex capable of activating FcɛRI, predictably due to the inadequately large distances between the sites in the oligomeric antigen bound by several OE-2. In contrast, OVA-resin is a multivalent antigen that carries many OVA with various distances. Thus, OVA-resin formed immune complexes with OE-2 containing many OVA/OE-2 complexes, some of which may have Fc portions that generate acceptable distances for FcɛRI bridging, resulting in degranulation responses. The characteristics of immune complexes differed among the composed antibodies and antigens, in turn, recognized epitopes. Thus, differences in the recognizing epitopes of OE-1 and OE-2 may be one of the reasons why OE-1 activates FcɛRI, whereas OE-2 does not. It is important to note that, based on the multiple mechanisms underlying the activation of FcɛRI by IgE, differences other than the epitopes recognized by these two antibodies may also be involved due to their distinctive competences in the activation of FcɛRI. Two mechanisms, the induction of activating signals from FcɛRI by monomeric SPE-7 without the antigen(24) and monomeric OE-1 with OVA mainly constructed by monomeric and dimeric forms (Yamaki et al., manuscript submitted), suggest that crosslinking is not the sole determinant of the stimulation of FcɛRI. Consistent with these findings, OE-2 only weakly induced FcɛRI-mediated activation of RBL2H3 cells even after stimulation with a multivalent antigen, namely, OVA-conjugated agarose. An analysis of conformational changes in OE-2 that induce activation of FcɛRI upon appropriate stimulation may provide important information on the other unknown domains of IgE responsible for its ability to stimulate FcɛRI.
Although the impact of glycosylation of IgE on its binding activity to FcɛRI and FcɛRII remains controversial, the deglycosylation of IgE appears to diminish its ability to bind to FcɛRI,(33) whereas IgE binds to FcɛRII regardless of the glycosylation level of the antibody.(34) Since OE-1 and OE-2 exhibited similar binding ratios for FcɛRI/FcɛRII in the present study, variations in the glycosylation of IgE may not explain why OE-2 only induced FcɛRII-dependent activation and not FcɛRI-dependent activation. A comparison of the functions of IgE carrying various glycosylation patterns may resolve this issue. Deglycosylated IgE at asparagine-394, which binds to the antigen, but not to FcɛRI,(33) has potential as an FcɛRII-specific activator, similar to OE-2. However, it is important to note that OE-2 binds to, but does not activate cells with FcɛRI in contrast to deglycosylated IgE at asparagine-394.
The limitation of this experiment was that we were unable to use class-switch variants, which have identical specificities for antigens but differ in constant regions. These variants may provide more accurate results, thereby clarifying differences in the functions of these antibodies. Nevertheless, the establishment of hybridoma producing the unique IgE antibody OE-2 in the present study was meaningful. The DNA sequence of this unique antibody will be available. Class-switch variants may only be produced after sequencing. This type of antibody had not been produced prior to the establishment of OE-2, because many monoclonal IgE already created, other than OE-2, did not activate FcɛRII without affecting FcɛRI activity. The series of IgE composed of various proportions of OE-1 and OE-2 made by genetic engineering may accelerate investigations on important portions in IgE and epitopes in OVA for the activation of FcɛRI and FcɛRII. We also cannot exclude the possibility that other putative IgE receptors, FcγRIV and/or galectin-3 in addition to FcɛRII, mediated the release of NO from macrophages and enhanced antibody responses in mice by OE-1 and OE-2. The production of tumor necrosis factor-α by mouse peritoneal macrophages stimulated with an IgE immune complex was previously shown to be dependent on FcγRIV.(7) Further studies are needed in order to elucidate the roles of these receptors in IgE-induced responses.
In conclusion, a newly produced anti-OVA monoclonal IgE, OE-2, had the unique ability to induce FcɛRII-dependent activation in the presence of the recognized antigen without inducing FcɛRI-dependent activation. The use of this antibody allows for FcɛRI- and FcɛRII-mediated functions of IgE to be easily distinguished. The availability of OE-2 in addition to conventional IgE, including OE-1, SPE-7, and 2C6, will advance research on the molecular mechanisms underlying the activation of FcɛRI and FcɛRII by IgE with its distinctive characteristics.
Footnotes
Acknowledgments
The authors thank Medical English Service (Kyoto, Japan) for editing the manuscript. Author Kouya Yamaki planned and performed all the experiments described herein and drafted the manuscript, and Shin Yoshino supervised the laboratory and assisted in the preparation and proofing of this article.
Author Disclosure Statement
The authors have no financial interests to disclose.