
Abstract
Purification of immunoglobulin M (IgM) antibodies could be challenging, and is often characterized by the optimization of the purification protocol to best suit the particular features of the molecule. Here, two different schemes were compared to purify, from ascites, the 1E4 IgM monoclonal antibody (mAb) previously raised against the stage of redia of the trematode Fasciola hepatica. This immunoglobulin is used as capture antibody in an immunoenzymatic assay to detect parasite ongoing infection in its intermediate hosts. The first purification protocol of the 1E4 mAb involved two chromatographic steps: an affinity chromatography on a Concanavalin A matrix followed by size exclusion chromatography. An immunoaffinity chromatography was selected as the second protocol for one-step purification of the antibody using the crude extract of adult parasites coupled to a commercial matrix. Immunoreactivity of the fractions during purification schemes was assessed by indirect immunoenzymatic assays against the crude extract of F. hepatica rediae, while purity was estimated by protein electrophoresis. Losses on the recovery of the antibody isolated by the first purification protocol occurred due to protein precipitation during the concentration of the sample and to low resolution of the size exclusion molecular chromatography step regarding this particular immunoglobulin. The immunoaffinity chromatography using F. hepatica antigens as ligands proved to be the most suitable protocol yielding a pure and immunoreactive antibody. The purification protocols used are discussed regarding efficiency and difficulties.
Introduction
S
Most of the developed mAbs are immunoglobulin G (IgG) isotype, while generation of other isotypes such as IgM is less common. Accordingly, several simple and efficient procedures for IgG purification have been devised and are widely available (e.g., ion exchange or affinity chromatography using general ligand such as Protein A and Protein G).(2,3) However, when dealing with parasites helminthes, the generation of IgM mAbs is not unusual.(4–6) The purification of this type of immunoglobulin could be tricky, since the size of this pentameric molecule generally leads to lower solubility, which increases sensitivity to denaturalization and decreases conductivity. These features could limit the application of standard purification tools. High molecular weight is related to slower diffusion constant that occasionally might hinder the use of size exclusion. Low conductivity can limit the application of ion exchange chromatography, while lower solubility and stability at extremes of pH could be problematic for affinity chromatography.(3) Therefore, the purification of each IgM requires optimization regarding the characteristics of the particular antibody.
Four anti-Fasciola hepatica rediae mAbs were developed for immunodiagnosis purposes and they recognize different antigenic preparations from several stages of the life cycle of the parasite, including the excretion–secretion antigens of F. hepatica adults. A double mAb-based immunoenzymatic assay (enzyme-linked immunosorbent assay [ELISA]) was developed and proved to be successful in the detection of the parasite inside its intermediate hosts when using the IgM mAb 1E4 as antigen-capturing antibody.(7) The high values of sensitivity and specificity of the developed ELISA assessed on laboratory-reared snails led the way to further studies for field validation of this immunoassay as a novel tool for the surveillance of fasciolosis transmission. This constitutes a challenge for the authors' laboratory, since the production of sufficient amounts of quality immunological reagents (i.e., the antibodies) is mandatory.
So far, the purification of the 1E4 mAb from mice ascites has been carried on by a two-step protocol using an affinity chromatography on a Concanavalin A Sepharose (Con A) column and a size exclusion chromatography on a Superose 6 column as the final step. This protocol is time-consuming, and often losses in antibody yielding have occurred during concentration by ultrafiltration prior to the size exclusion procedure. Therefore, the aim was to devise a one-step purification approach based on immunoaffinity chromatography for the purification of 1E4 mAb from ascites and to compare it with the former protocol to select the most suitable methodology. Purity, immunoreactivity, antibody recovery, as well as the time and the cost of the process were the considered variables.
Materials and Methods
F. hepatica and preparation of crude extracts
Adult parasites of F. hepatica species were obtained in cattle abattoirs. The trematodes were transported to the laboratory in saline solution (0.85% NaCl) supplemented with 5% glucose for egg laying. Then, the parasites were thoroughly washed with saline solution, and a total extract of F. hepatica adults was obtained in cold 0.1 mol/L carbonate-bicarbonate buffer pH 8.8, using a Polytron® laboratory homogenizer (Kinematica, Luzern, Switzerland). The homogenate was clarified by centrifugation at 10,000 g, 4°C, for 15 min, and protein concentration was quantified by the bicinconinic acid reaction.
F. hepatica rediae were obtained from intermediate host snails that were experimentally infected with the parasite.(8) A crude extract of F. hepatica rediae was obtained according to the methodology described by Alba et al.(9)
In vivo production of 1E4 monoclonal antibody
For the production of 1E4 mAb, BALB/c mice supplied by the National Center for Production of Laboratory Animals of Cuba were used. Animal welfare was guaranteed, and all procedures involving experimental animals were approved by the Institutional Animal Care and Use Committee of the Institute “Pedro Kourí” of Tropical Medicine. 1E4 mAb-producing hybridoma was inoculated into the peritoneal cavity of mice, and ascites was produced and collected from euthanized animals. An indirect ELISA using Maxisorp (Nunc A/S, Roskilde, Denmark) microtiter plates coated with 10 μg/mL of the crude extract of F. hepatica rediae diluted in carbonate-bicarbonate buffer pH 9.6(7) was used for assessing the immunoreactivity of the ascites and of the different fractions derived from the purification procedures.
First purification protocol involving two chromatographic steps
1E4 mAb was partially purified from ascites by chromatography on Concanavalin A Sepharose Fast Flow column (GE Healthcare Life Sciences, Pittsburgh, PA) using a Fast Protein Liquid Chromatography system (FPLC; Pharmacia, Uppsala, Sweden) and 1 mL/min of flow rate. The matrix was equilibrated with binding buffer (20 mmol/L Tris-HCl, 0.5 mol/L NaCl, 1 mmol/L CaCl2, pH 7.5), and ascites was applied onto the column previously diluted 1:2 in the binding buffer. The matrix was washed, and the unbounded fraction of the ascites was collected. The matrix-bounded fraction was eluted using the binding buffer with 0.1 mol/L α-methyl-D-mannoside (Sigma, St Louis, MO) and dialyzed against phosphate-buffered saline (PBS), for 16 h at 4°C. Afterwards, the eluted fraction was concentrated in AMICON® devices using YM-100 membranes (Millipore, Bedford, MA) and fractionated on Superose 6 prep grade matrix (GE Healthcare Life Sciences) packed onto a 16 × 60 column. A maximum of 1 mL of sample per chromatography was applied, and PBS was used as running buffer at a flow rate of 0.5 mL/min.
Second purification protocol: immunoaffinity chromatography
To select the antigenic mixture for devising the immunoaffinity matrix, the immunoreactivity of the 1E4 mAb (purified by the first protocol) against the total extract of F. hepatica adults was compared with the immunoreactivity against the crude extract of F. hepatica rediae in an indirect ELISA.(7) Microtiter plates sensitized with 10 μg/mL of each antigen were used. Then, an affinity column was prepared by immobilizing 150 mg of proteins of the total extract of F. hepatica adults to 10 mL of glutaraldehyde-activated α-amino hexyl sepharose 4B (Sigma, St Louis, MO), according to the manufacturer's directions.
The immunoaffinity chromatography was performed in a FPLC system using a flow rate of 1 mL/min. The immunoaffinity matrix was equilibrated with PBS, and 1E4 mAb-containing ascites was applied to the column diluted 1:2 in PBS. The unbounded fraction was collected until base line was recorded. Then, the ascites fraction captured by the matrix was eluted with 0.1 mol/L glycine-HCl buffer, pH 2.8, into neutralization buffer (1 mol/L Tris-HCl, pH 8). Two immunopurification cycles per sample were performed, and the eluted fractions were pooled and dialyzed in PBS for 16 h at 4°C.
Assessment of purification parameters
Protein concentration and immunoreactivity against the crude extract of F. hepatica rediae of all chromatographic fractions (derived from both purification protocols) were determined by the bicinconinic acid reaction and indirect ELISAs, respectively. Then, all immunoreactive fractions were kept with 0.01% sodium azide at 4°C, and the 1E4 mAb recovery of both purification schemes was calculated in milligrams of purified mAb per milliliter of ascites.
The purity of the final 1E4 mAb-containing fractions isolated with both purification protocols was analyzed on a 12% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE).(10) The samples were diluted with 2-mercaptoethanol as reducing reagent, and 0.1% Coomassie Brilliant Blue R-250 (Merck, Darmstadt, Germany) was used for gel staining. The stained gel was analyzed by scanning densitometry using ImageJ software v1.41, and the purity of the isolated 1E4 mAb was estimated.
Statistical analysis
Mean and standard deviation for triplicate values of optical density obtained with the indirect ELISAs were calculated. Data were analyzed by the Shapiro–Wilk statistical test to verify normal distribution, whereas Levene's test was used to check the variance homogeneity. A Mann–Whitney U test was then performed to assess statistical differences regarding the immunoreactivity of 1E4 mAb, used as control (purified by the Con A–size exclusion protocol), against the total extracts of F. hepatica adults and of rediae in order to select the antigenic mixture for devising the immunoaffinity matrix. This test was also used to compare the immunoreactivity against the crude extract of F. hepatica rediae between the two sets of 1E4 mAb isolated by each of the purification protocols. Statistica software v7.0 was used for the analysis, and differences were considered significant when p < 0.05.
Results and Discussion
The purification of IgM antibodies is often characterized by the optimization of the purification protocol to best suit the particular features of the molecules, and is usually characterized by a combination of different methods.(1) To select a particular purification protocol, several factors must be taken into account: purity, biological activity, recovery of the antibody, as well as the time and cost of the process.(3) All these aspects were considered in order to choose the most suitable methodology.
The first purification scheme included two different principles of antibody purification: affinity for a general ligand, and size exclusion relying on the molecular weight of the molecule. The partial purification of a highly reactive anti-F. hepatica rediae ascites by a Con A ligand showed a classical pattern of two fractions (unbounded and eluted) of affinity chromatography (Fig. 1A). The unbounded fraction showed no recognition of the crude extract of rediae by indirect ELISA, which suggests that all the 1E4 mAb was efficiently captured by the matrix. The affinity chromatography using specific glycan-binding lectins (e.g., Con A) has been used for IgM purification due its high number of glycosylated sites.(11–13) However, other proteins of ascites apart from IgMs also have glycan chains, which can interact with the matrix. Therefore, a further purification step is often required. In this sense, the size exclusion chromatography is suitable, since the high molecular weight of IgM allows its separation from most of the remaining mixtures of proteins by this principle.(2) The gentle separation condition of this chromatography overcomes possible loss of the antibody's immunoreactivity that occasionally occur when other methods with different requirements (i.e., exposure to extreme pH, high concentration of salt, chaotropical agents) are applied.(3)
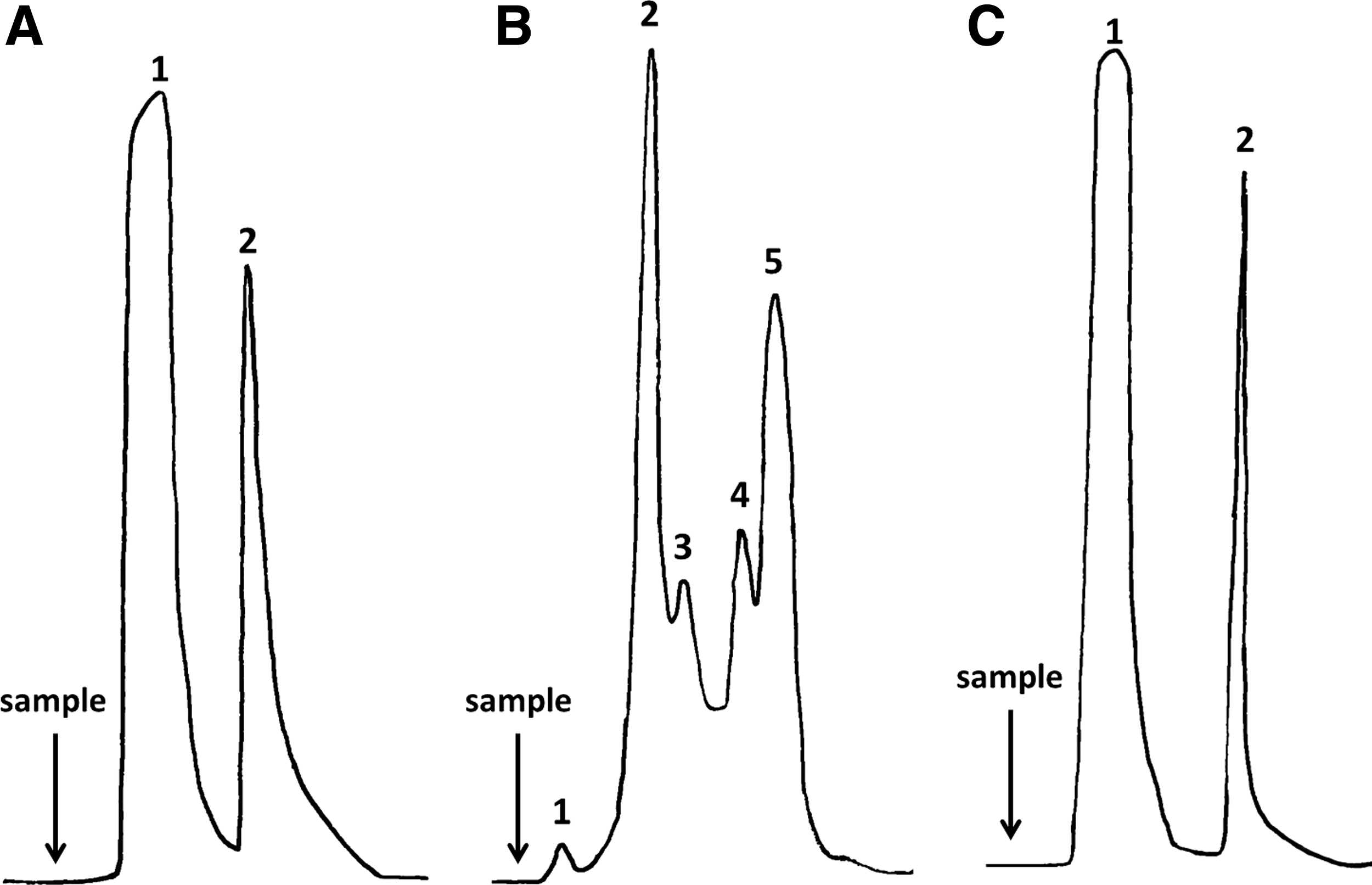
(
Figure 1B shows the profile of the size exclusion chromatography of the Con A-eluted fraction. The small peak eluted within the void volume of the column (Peak 1) is most likely related to aggregates of high molecular weight that probably occurred during the concentration step that followed the Con A chromatography. This fraction has been proven to be highly unstable, since precipitation has been recorded later on. Noteworthy, immunoreactivity against the crude extract of F. hepatica rediae was confirmed in Peak 2 and less in Peaks 3 and 4. This result is likely associated with the delay of the mAb during the chromatography and thus the mixing of part of it with proteins of lower molecular weights (in fact, no baseline occurred between these fractions). This behavior could be related to a slow diffusion constant of the 1E4 mAb that might be associated with its large size, which decreases the resolution of a method that relies on diffusion for mass transport. However, Peak 2 showed the highest immunoreactivity against the crude extract of F. hepatica rediae as well as the highest intensity, which is directly related to the amount of high molecular weight proteins (such as IgM antibodies). Accordingly, Peak 2 was identified as the purified 1E4 mAb-corresponding fraction and was further analyzed by SDS-PAGE to assess purity.
Antigen-specific antibodies can be obtained by immobilizing a specific antigen on a matrix and using it for isolating antibodies. This type of immunoaffinity chromatography exploits the specificity of binding mAb-cognate antigen for selective absorption of the particular antibody from a complex mixture, and can be a very powerful, rapid, and highly specific procedure.(14) Therefore, this approach was selected as a second purification protocol to isolate the 1E4 mAb from ascites using a one-step procedure.
Since the availability of the antigen that needs to be immobilized on the resin is usually a major drawback of specific ligand affinity chromatography,(3) this issue was analyzed when devising the protocol. In this sense, the recombinant expression of the antigen and the chemical synthesis of ligand peptides are feasible alternatives for large-scale production of pure antigens. However, it is only possible to apply them when the antigen is well characterized. Instead, purification of the specific antigen from complex mixture is time-consuming, and productivity could be an issue to consider. Therefore, an immunoaffinity matrix was devised using a complex mixture of F. hepatica antigens that was selected taking into account the suitability of the developmental stages of the parasite recognized by the 1E4 mAb. In this sense, while the production of redia larvae is difficult and time-consuming due to the need to perform experimental infections of intermediate hosts followed by snail dissection, the collection of adult parasites in abattoirs is relatively simple and low cost. In addition, higher amounts of protein are obtained from a single F. hepatica adult compared with rediae. The representation of the epitope on both extracts was also considered for the selection of the antigenic preparation to be immobilized in the matrix. Since similar results (p = 0.31232) were assessed regarding the reactivity of the purified 1E4 mAb against the total extract of F. hepatica adults (1.228 ± 0.079) and the crude extract of rediae (1.415 ± 0.1369), the former antigenic preparation was judged as convenient to use to devise the immunoaffinity matrix.
Figure 1C shows the profile of 1E4 mAb-containing ascites fractionated with the immunoaffinity chromatography. The unbounded fraction did not show recognition of the crude extract of F. hepatica rediae, while the eluted fraction displayed a high reactivity against this antigen. In addition, the immunoreactivity of the eluted fraction against the crude extract of F. hepatica was similar to that of the 1E4 mAb purified by the first protocol (p = 0.1912). This result indicates that the specific antibody was captured by the total extract of F. hepatica adults immobilized on the matrix, and then it was completely eluted without losing its reactivity against the crude extract of F. hepatica rediae. Noteworthy, no precipitation occurs during this protocol, and the antibody displayed resistance to pH change, since no affectation on the recognition of larval antigens of the parasite was assessed. Therefore, concerns regarding possible denaturalization of the mAb and loss in immunoreactivity during elusion were set aside.
The comparison regarding 1E4 mAb yielding between three cycles of purification with both protocols resulted in an average recovery of 0.84 mg/mL of ascites with the first purification scheme in contrast to a yield of 1.26 mg/mL of ascites achieved with the second protocol. This difference is likely related to protein precipitation assessed during the concentration of the Con A-eluted fraction and to the resolution of the size exclusion chromatography regarding this particular antibody discussed previously. All of these events ultimately affect the recovery of the antibody that is in addition highly diluted after the size exclusion chromatography. For purity analysis, the 1E4 mAb-containing fractions isolated with both protocols were resolved on a reducing SDS-PAGE (Fig. 2). The purity of the antibody was estimated and resulted in 71.7% with the first purification protocol, while a higher purity of 83.47% was achieved with the immunoaffinity chromatography. Matrix specificity and binding capacity was checked after each of six different purification cycles of the 1E4 mAb. It was noticed that the same matrix could be reused in four different occasions to isolate the antibody without any significant loss regarding its immunoreactivity, recovery, or purity.

Sodium dodecyl sulfate polyacrylamide gel electrophoresis on 12% polyacrylamide gel. Lane 1: molecular weight standard from 10 to 225 kDa; lane 2: 1E4 mAb purified by the first purification protocol (Con A–size exclusion protocol); lane 3: 1E4 mAb purified by the second purification protocol (immunoaffinity chromatography). Coomassie Brilliant Blue (0.01%) solution was used for protein staining.
To conclude, these results suggest that immunoaffinity chromatography could be a suitable way to purify 1E4 mAb with a time-saving and low-cost scheme. Further studies regarding the characterization of the particular epitope recognized by the 1E4 mAb would be helpful in order to improve the capacity of the immunoaffinity column through the substitution of the crude extract of F. hepatica adults for specific synthetic peptides on the matrix.
Footnotes
Author Disclosure Statement
The authors have no financial interests to disclose.