
Abstract
CD275 (B7-H2, ICOSL), a co-stimulatory molecule of the B7 superfamily, plays a critical role in immune response. In this report, a novel mouse anti-human CD275 monoclonal antibody (MAb) was prepared using hybridoma technology, and immunological characteristics of the MAb were determined. The results showed that the MAb (clone 13D11) was IgG2(κ) and bound specifically to human CD275. By mutual competition, we found that the antibody recognized different epitopes of CD275 antigen compared with commercial antibodies and could block ICOS-CD275 interaction. Crosslinking of CD275 with MAb 13D11 markedly blocked ICOS positive signal and inhibited T cell proliferation and cytokine production. In addition, the 13D11 MAb was suitable for indirect ELISA detection. Thus, the MAb against human CD275 with high specificity and different activity would be useful for further study of this molecule.
Introduction
CD275,
CD275 is expressed by a wide variety of cell types, including B cells, macrophages, dendritic cells (DCs), and a subset of T cells,(5–7) and can be induced on epithelial cells, vascular endothelial cells (ECs), and fibroblasts, as well as in many primary tumors and tumor cell lines.(8,9) Recently, Maazi and colleagues showed that ICOS-CD275 interaction promoted cytokine production and survival in Type 2 innate lymphoid cells (ILC2s) through STAT5 signaling.(10) Dianzani and colleagues found that CD275 triggering strikingly inhibited the migration activity of both tumor and ECs in vitro, which was associated with the dephosphorylation of focal adhesion kinase (FAK) and decreased β-Pix expression.(11) Previously, we found elevated expression of CD275 on thyroid follicular cells (TFCs) from GD patients, and demonstrated that ICOS-CD275 signal plays a direct role in proliferation and differentiation of TFC and may exert important effects in the initiation, maintenance, and exaggeration of autoimmune responses in local tissue.(12) Blocking the function of CD275 on B cells by antibodies or via conditional genetic deletion abrogates GC formation.(13,14) In summary, identification of CD275 in different pathological conditions suggests that it may become an interesting target for disease diagnosis or therapy.
Here we describe the development of the CD275-specific antibody. We have successfully produced and characterized a novel mouse monoclonal anti-human CD275 antibody based on flow cytometry, dot-blot and Western blot analyses, enzyme-linked immunosorbent assay, immunofluorescence staining, and functional in vitro assays. This antibody provides a useful tool to evaluate the effect and measure the level of CD275 in numerous biological processes.
Materials and Methods
Cell lines and reagents
Mouse fibroblast L929 cells were stably transfected with empty vector (L929/mock cells), human CD275 (L929/CD275 cells), or B7-H4 genes (L929/B7-H4 cells), as described previously.(15) RPMI-1640 media were purchased from Gibco (Carlsbad, CA). All the cells were cultured in RPMI-1640 medium supplemented with 10% fetal calf serum (FCS, Sijiqing Co., Hangzhou, China). PE-conjugated mouse anti-human CD275 (clone no. 2D3) antibody was purchased from Biolegend (San Diego, CA). Functional CD275 antibody (clone no. MIH12) was the product of eBioscience (San Diego, CA). Commercial recombinant ICOSIg protein was obtained from R&D Systems (Minneapolis, MN). Biotinylated goat anti-mouse IgG1, PE-conjugated goat anti-mouse IgG, mouse IgG1 (clone 679.1Mc7), and mouse anti-human CD3 MAb (clone UCHT1), were all products from Immunotech (Marseille, France). HAT and HT products were purchased from Sigma (St. Louis, MO). BD CBA Human Th1/Th2/Th17 Cytokine Kit and IgG isotyping kit were from BD Biosciences (San Jose, CA).
Generation of anti-human CD275 MAbs
L929/CD275 cells were used as immunogens. BALB/c mice (female, 6–8 weeks old) were chosen and intraperitoneally injected with L929/CD275 cells pretreated by mitomycin. According to the method described by Groth and Scheidegger, the booster injection was repeated three times every 3 weeks.(16) Then the splenocytes and mouse myeloma cells (SP2/0 cell line) were fused in the presence of 50% polyethylene glycol (PEG) 3 days later. The fusion cells were cultured with HAT-DMEM medium containing 15% FCS in 96-well plates to select hybrid clones. All hybrid clone supernatants were screened for the detection of antibody using flow cytometry. The hybrid clone secreting antibody that was reactive with L929/CD275 cells but not with L929/mock cells or L929/B7-H4 cells was selected and subcloned by limiting dilution method three times to establish hybridoma cell line secreting monoclonal antibody. Mouse MAbs were produced from female BALB/c mice in which hybridoma cells had been injected intraperitoneally into pristane-primed BALB/c mice for 8–10 days. The MAbs were purified from the mouse ascites on Protein G Sepharose affinity column.
Characterization of MAbs
Ig isotypes were identified with the test paper (Roche, Mannheim, Germany), and CBA isotyping kit (BD Biosciences), according to the manufacturer's instructions. The mutual competition assay was performed to determine whether the two antibodies recognize the same or overlapping epitopes of human CD275 antigen. Thus, the competition of all possible pairs of biotinylated and unlabeled anti-CD275 MAbs were assessed by flow cytometry. L929/CD275 cells were incubated with one of the MAbs (ranging from 0.01 to 10 μg) used as competitors for 30 min at 4°C. After being washed with PBS, the other biotinylated MAbs (1 μg/test) were added respectively for another 30 min, followed by streptavidin-PE for 30 min. The fluorescence intensity of stained cells was analyzed by flow cytometry. Mutual competition assay between these anti-CD275 MAbs and the recombinant ICOSIg proteins was also used to assess the spatial impact of the MAbs to ICOS-CD275 interaction.
Dot-blot and Western blot analyses
Dot-blot and Western blotting assays were used to analyze the binding ability of the two MAbs to recombinant CD275-Ig. CD275-Ig was loaded in equal amounts on nitrocellulose membranes or separated by SDS-PAGE, followed by immunoblotting with MAb produced by hybridoma clones for CD275. Briefly, samples were mixed with Laemmli buffer, boiled at 95°C for10 min, and loaded onto SDS-PAGE. Proteins were separated by electrophoresis and blotted onto nitrocellulose (Pierce, Rockford, IL). Non-specific binding was reduced by blocking the membrane in 5% non-fat dry milk in PBS (pH 7.2) on a rocking platform. The membrane was then washed with PBS containing 0.1% Tween-20 (PBST), and incubated with anti-CD275 MAbs (13D11 or 2D3) or mouse IgG1 for 2 h at RT. Then the membrane was incubated with horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG for 1 h. After four washes, enhanced chemiluminescence (ECL, Pierce) was applied to the membranes, which were then exposed to an X-ray film (Kodak, Rochester, NY).
Enzyme-linked immunosorbent assay of MAbs
The application of the obtained MAbs was determined by indirect ELISA. Briefly, the recombinant CD275-Ig fusion protein diluted to different concentrations (5, 1.25, and 0.3125 ng/mL) was added to each well of an ELISA plate and incubated at 4°C overnight. After extensive washing with PBS containing 0.1% Tween-20, every well was blocked with 2% bovine serum albumin (BSA) for 1 h at 37°C. Then the obtained anti-CD275 MAbs (1 μg/mL) were added for 2 h at room temperature, followed by HRP-goat anti-mouse IgG at 1:2000 for 1 h. Color development was performed by adding 100 mL of 0.1 mg/mL tetramethylbenzidine in 0.1 M citrate-phosphate buffer (pH 5.0) containing 0.01% H2O2. The absorbance at 450 nm was determined with a microplate reader (Bio-Rad, Hercules, CA).
Immunofluorescence microscopy
L929/CD275 cells were grown on cover glasses in RPMI 1640 medium supplemented with 10% FBS for 24 h. The cells were labeled with MAbs (1:200) for 1 h. After three 10 min washes with TBST buffer, the cells were incubated with goat anti-mouse secondary antibodies conjugated to Alexa Flour 594 (BD Biosciences). Subcellular localization of CD275 was viewed under a confocal fluorescent microscope.
Analyzing proliferative and cytokine responses
In vitro proliferation assays based on CFSE dilution were performed. Briefly, PBMCs were isolated from healthy controls, and cells 1 to 10 million were resuspended in 2 mL CFSE at the desired dilution and incubated at 37°C for 10 min. 1 × 106 cells were stimulated with CD3 (100 ng/mL) in the presence or absence of 13D11 (0 ∼ 10 ng/mL) at 37°C, 5% CO2. After 6 days in culture, the cells were stained for surface markers with Alexis Fluor 700-conjugated anti-CD3 (clone SP34-2; BD Biosciences). Supernatants from similar cultures were collected after 6 days for assessment of cytokine production using a Cytometric Bead Array (CBA) (BD Biosciences).
Cytometric bead array
The culture supernatants were collected, and cytokine levels were measured by Human Th1/Th2/Th17 CBA kit (BD Biosciences). Briefly, 50 μL of samples or known concentrations of standard samples were added to a mixture of 50 μL capture bead and 50 μL of phycoerythrin-conjugated detection antibody mixture for 2 h at room temperature in the dark, and then washed to remove unbound detection antibody. Data were acquired using a FACS Aria II Flow cytometer and analyzed using CBA software 1.1 (BD Biosciences).
Statistical analysis
All statistical analyses were performed using GraphPad Instat (v. 3.10), and the graphs were plotted using GraphPad Prism (v. 5.03; GraphPad). For the comparison of two time points, the Mann-Whitney U test (two-tailed p value) was performed.
Results
Generation of five mouse anti-human CD275 MAbs
Human CD275 gene was cloned and expressed constitutively on the membrane of L929/CD275 transfectant (Fig. 1A). Five mouse anti-human CD275 MAbs (clones 20B10, 6D7, 7B9, 16B8, and 13D11) were obtained in this study using L929/CD275 cell line as the immunizing antigen in BALB/c mice. The specificity of the MAbs for human CD275 was first identified by flow cytometry. They could bind to L929/CD275 transfected cells but not to L929/mock and L929/B7-H4 transfectants (Fig. 1B). MAbs 20B10, 7B9, and 13D11 were IgG2(κ), 6D7 was IgG3 κ, and 16B8 was IgMκ (Fig. 2). 13D11 MAb was assigned for detailed characteristic assay. With recombinant CD275-Ig protein as antigens, the results of dot-blotting and Western blotting showed that 13D11 could recognize the target protein bands (Fig. 3A, B). The MAb was also identified by ELISA. The results indicated that 13D11 can bind to immobilized CD275-Ig protein in indirect ELISA (Fig. 3C). Meanwhile, the results indicated that 13D11 could recognize CD275 protein in immunofluorescence microscopy (Fig. 3D). Thus, the novel MAb 13D11 against human CD275 reported in this work provides additional reagents for the expression and functional characterization of CD275.
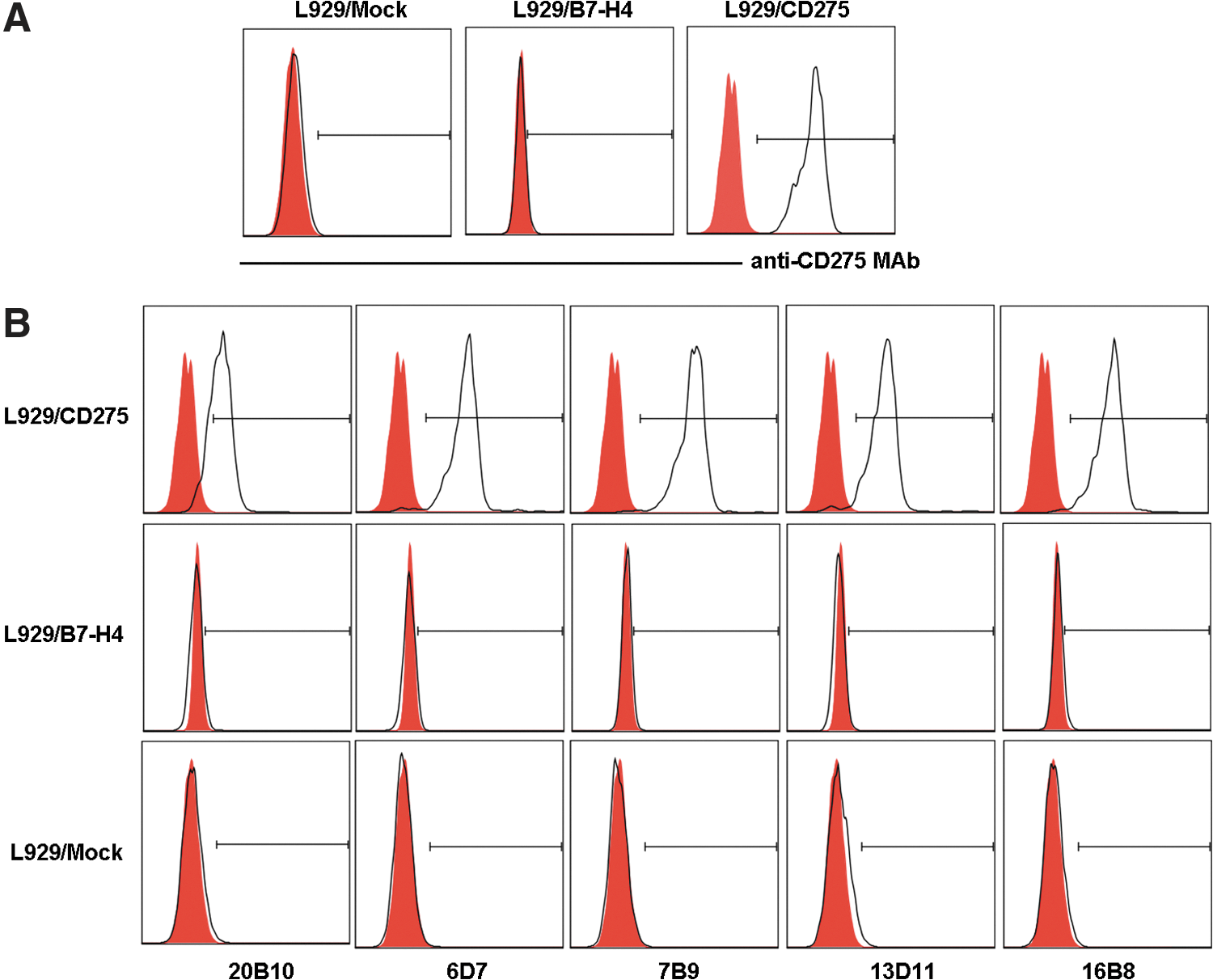
Generation of five MAbs against human CD275. (
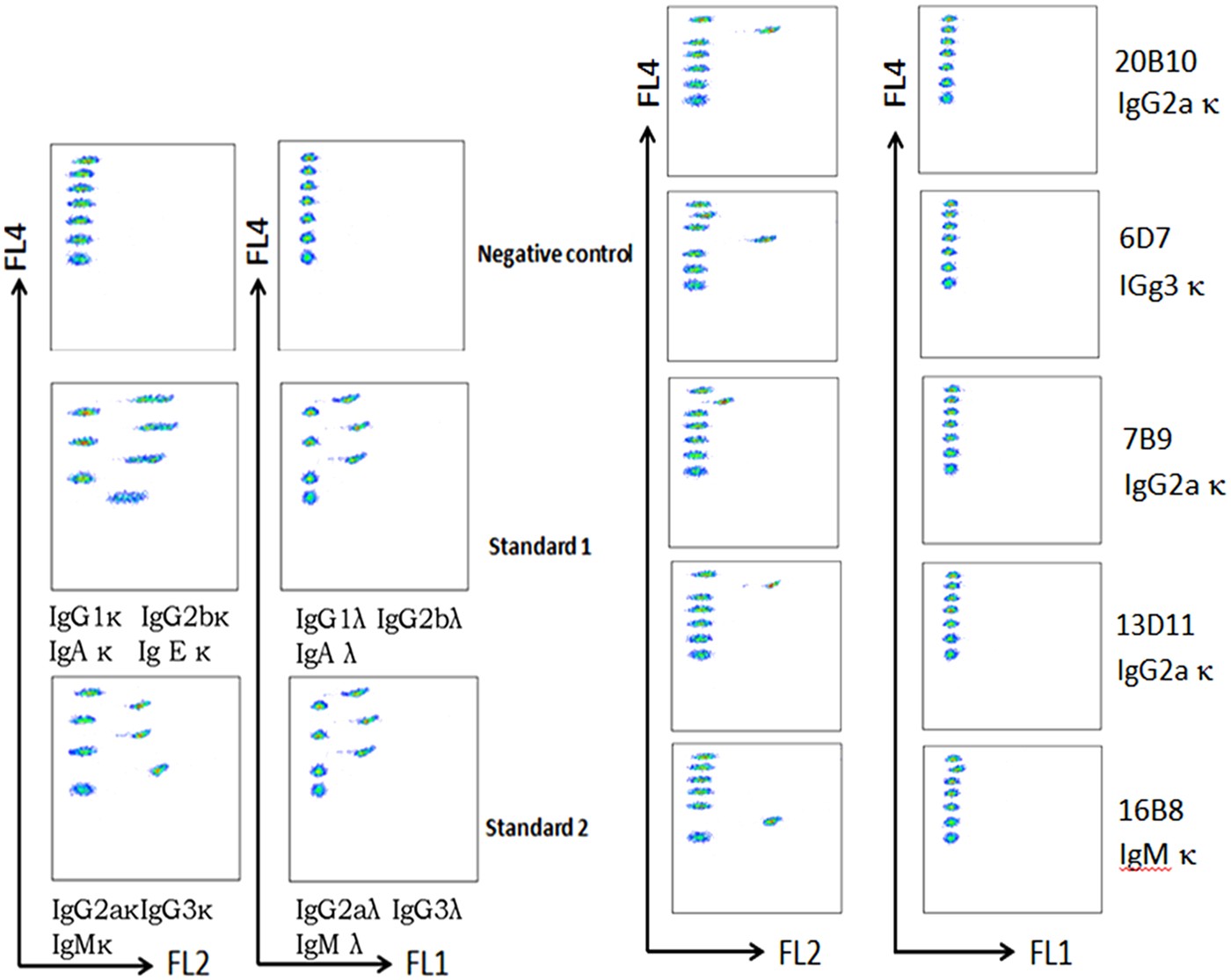
Isotype of mouse monoclonal antibody against human CD275. Isotypes of MAbs were determined using a mouse monoclonal isotyping kit by CBA method according to the manufacturer's instructions. CBA, cytometric bead array.

Characterization of MAbs. (
Epitope recognized by 13D11 is different from that of commercially available 2D3 MAb
In order to determine whether the new antibody recognizes different epitopes, mutual competition assay was performed by flow cytometry. In this work, the binding of one biotin-conjugated MAb to L929/CD275 cells was competed by other unlabeled MAbs. As shown in Figure 4, graded amounts of unlabeled 13D11 blocked subsequent binding of biotinylated MAb to L929/CD275cells in a dose-dependent way and 0.5 μg was sufficient for both antibodies. The data indicate that 13D11, 2D3, and MIH12 MAbs recognize three different epitopes of human CD275.
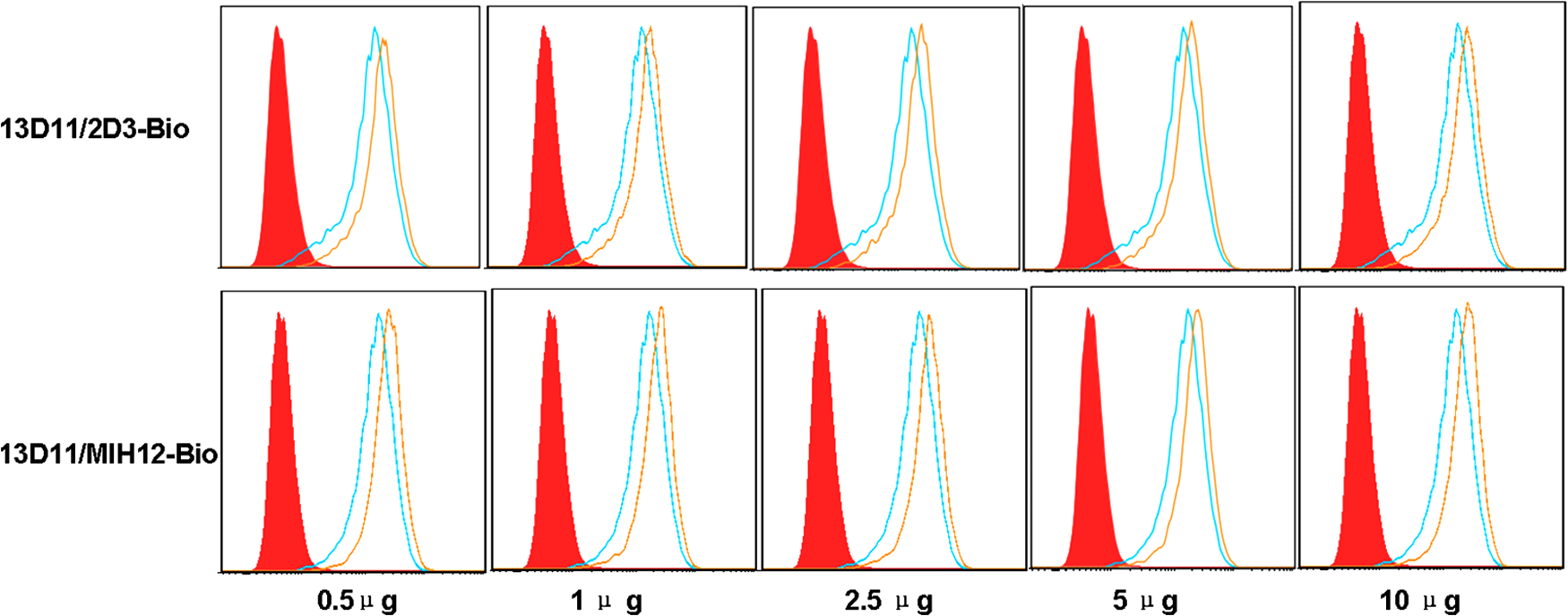
Mutual competition assay of MAbs. L929/CD275 cells were incubated with saturating concentration of one unlabeled MAb for 30 min at 4°C; then cells were washed and stained with other biotinylated anti-CD275 MAbs (blue lines) followed by streptavidin-PE for 30 min at 4°C. Fluorescence intensity of stained cells was analyzed by flow cytometry. L929/CD275 cells were also stained directly with biotinylated mouse IgG1 followed by streptavidin-PE as negative control (red solid histograms) or biotinylated anti-CD275 MAbs as positive control (yellow lines). The figure is representative of three experiments.
MAb 13D11 blocks ICOS-CD275 interaction
Mutual competition assay between the anti-CD275 MAbs and the recombinant ICOS-Ig proteins was also used to assess the spatial impact of the MAbs to ICOS-CD275 interaction. The results showed that the addition of different amounts of 2B4 and 13D11 MAbs could inhibit the binding of ICOS-Ig fusion protein to L929/CD275 cells (Fig. 5). These results suggest that the novel antibody 13D11 may be blocking antibodies for ICOS/CD275 co-stimulatory pathways.

Effect of anti-CD275 MAbs on ICOS-CD275 interaction. L929/CD275 cells were incubated with indicated doses of different MAbs for 30 min at 4°C; then cells were reacted with ICOS-Ig fusion protein followed by PE goat anti-human IgG(Fc) (yellow lines) for 45 min at 4°C. At the same time, L929/CD275 cells were stained directly with ICOS-Ig protein followed by PE-goat anti-human IgG(Fc) as positive control (blue lines) or human IgG as negative control (red solid histograms). Fluorescence intensity of stained cells was analyzed by flow cytometry.
Blocking MAb 13D11 inhibits T cell proliferation
To assess whether the blocking MAb 13D11 exhibited any functional effect on T cells, an in vitro direct proliferative assay was performed in the presence of 13D11 MAb PBMC cells. Compared with the control groups, the addition of 13D11 MAbs inhibits CD3+T cell proliferation in a dose-dependent manner, while 2B4 produced less inhibition (Fig. 6A). Meanwhile, co-culture of 13D11 with PBMC cells also inhibited IL-4, IL-10, TNF-α, and IFN-γ production by CD3+ T cells (Fig. 6B). Therefore, MAb 13D11 is a functional antibody that can inhibit both ICOS/CD275 co-stimulatory signals on T cells. These results demonstrate that 13D11 efficiently reacts with human CD275 and that 13D11 is the better MAb for blocking the interaction between CD275 and its receptor.
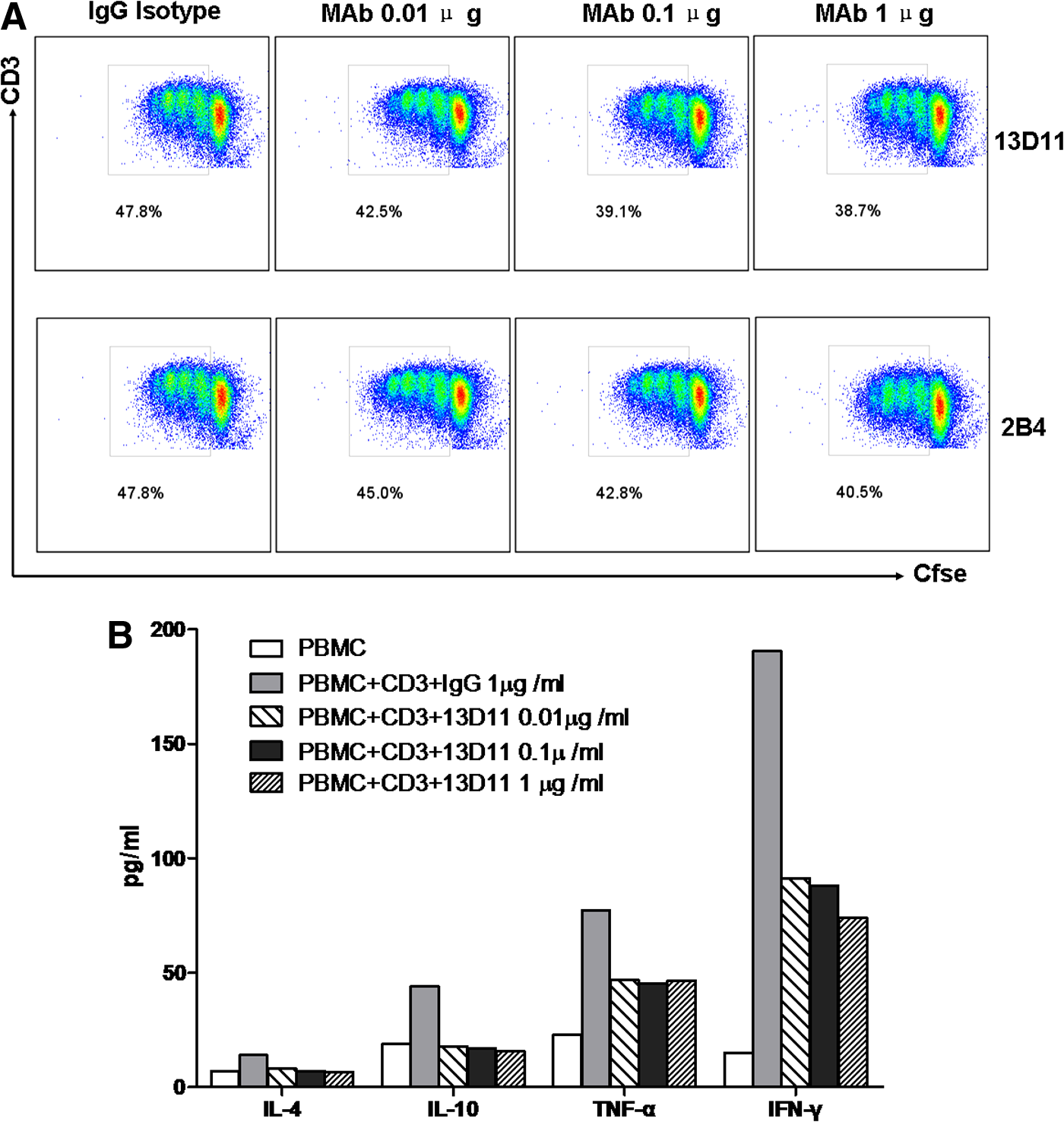
T cell proliferation upon engagement of CD275 with MAb 13D11. (
Discussion
Antibodies can specifically bind to its ligand and lead to target cell lysis or block the pathological process. Thus the development of antibodies in a particular humanized antibody drug plays an important role in clinical therapy of human malignancy and other autoimmune diseases. Our study describes the generation and characterization of a highly specific mouse monoclonal antibody against human CD275. This development is important for the further study of the role of CD275 in many biological processes.
The wide expression pattern of CD275 has made it an interesting target for therapy of many diseases. Recent reports have shown that the ICOS/CD275 signal pathway plays a crucial role in prevention of tumor development, as well as in the regulation of autoimmune diseases.(17–19) In this work, we produced an antibody against CD275 by using the L929/CD275 transfectant as immunogen to immunize BALB/c mice and successfully established a hybridoma cell line secreting MAb against CD275 molecule through hybridoma technology. Competitive inhibition experiment demonstrated that MAb 13D11 and commercial antibodies 2D3 and MIH12 bound to the different antigenic epitopes on the CD275 molecule. Moreover, mutual competition assay between the anti-CD275 MAbs and the recombinant ICOS-Ig proteins showed that 13D11 may bind (or nearly bind) to the ligand-binding site of CD275. These results suggest that the novel antibody 13D11 may be blocking antibodies for ICOS/CD275 co-stimulatory pathways.
It is well known that antibodies can specifically bind to its ligand and lead to target cell lysis or blockage of the pathological process. We found that 13D11 MAb can effectively inhibit T cell proliferation and decrease production of IL-4, IL-10, TNF-α, and IFN-γ, suggesting that this MAb may be useful for blocking the ICOS/CD275 positive signal, which closed up-regulation signal transduction by CD275 to T cell, and inhibited T cell reactivity to various antigens. As an inhibitor of CD275-mediated positive regulation of T cell activation, 13D11 MAb can inhibit the immune response. But the mechanism is still under research. CD275 and anti-CD275 MAbs, as important factors of T cell function, can play specific therapeutic effects on disease, which has high efficacy, through the intervention of an immune microenvironment. Thus we can open up a new pathway for gene therapy.
In conclusion, we developed a new MAb against CD275. This MAb had great utility in flow cytometry, immunoblotting, indirect immunofluorescence staining, and ELISA, and it also showed an inhibitive biological function to CD275. Accordingly, this monoclonal antibody will provide a powerful tool for further investigation of CD275 function.
Footnotes
Acknowledgments
This work was supported by the National Program on Key Basic Research Project (973 Program; grant no. 2013CB530501), the National Natural Science Foundation of China (grant nos. 31170834, 30930085), the Science and Technology Project of Suzhou (grant no. SYS201331), and the Pre-research Project of Suzhou University for National Natural Science Foundation (grant no. SDY2012B25).
Author Disclosure Statement
The authors have no financial interests to disclose.