
Abstract
NOB1, a transcription-associated protein, may play important roles in the development of many cancers. In this study, we have efficiently generated one monoclonal antibody (MAb) against NOB1. Enzyme-linked immunosorbent assay (ELISA) and western blot were used to screen the hybridomas. As a result, one MAb named L6 (IgG1) effective in detecting the recombinant and the cellular NOB1 protein was characterized. Using L6, NOB1 was found to be upregulated in gastric cancer cells and tissues compared with normal gastric epithelial cells and nonneoplastic tissues. The expression of NOB1 was also found to be higher in multidrug-resistant gastric cancer cells than that of sensitive cells. This novel MAb will be valuable for investigating the role of NOB1 in carcinogenesis and multidrug resistance of gastric cancer.
Introduction
T
Downregulation of NOB1 suppressed the proliferation and tumor growth of NSCLC in vitro and in vivo. Zhang et al. have found that knockdown of NOB1 expression could inhibit the malignant transformation of human prostate cancer cells.(6)
To understand its distribution in tissue as well as its function, establishing a monoclonal antibody (MAb) with specificity to NOB1 was necessary. In this study, we have produced and characterized a novel MAb specific to the NOB1. This MAb could recognize NOB1 protein derived from human cancer cell lines by western blotting.
Materials and Methods
Cell lines and tissues
The mouse myeloma cell line SP2/0, the normal gastric epithelial cell line GES, and human gastric cancer cell lines MKN28 and SGC7901 were routinely cultured in the RPMI 1640 medium (Life Technologies, Inc.) supplemented with 10% fetal calf serum in a 37°C humidified incubator with a mixture of 95% air and 5% CO2. Human vincristine-resistant gastric adenocarcinoma cell line SGC7901/VCR and adriamycin-resistant gastric adenocarcinoma cell line SGC7901/ADR were developed by exposing the parental SGC7901 cells to stepwise increasing concentrations of anticancer drugs (VCR or ADR) and kept in our laboratory.(7) To maintain the drug resistance phenotype of SGC7901/VCR cells and SGC7901/ADR cells, 1.0 μg/mL VCR and 0.8 μg/mL ADR were additionally added to the medium, respectively.
Five cases of gastric adenocarcinoma were chosen from gastric cancer patients operated on in 2014 in the Xijing Hospital, FMMU. None of the patients had received radiotherapy or chemotherapy before their operation and were affirmatively diagnosed by pathological analysis after the operation. Two sites were selected for protein extraction in each case: carcinoma tissue and adjacent nonneoplastic tissues.
Expression of NOB1 in Escherichia coli
Briefly, the total RNA extracted from SGC7901 cells was used for NOB1 cDNA synthesis. The PCR product was gel purified, ligated into pET28-a vector, and then transformed into Escherichia coli DH5. Clones that emerged on the plates containing kanamycin were analyzed by restriction endonuclease digestion. The correct recombinant plasmids were selected and transformed into competent E. coli BL21 (DE3). After induction of expression, the bacteria were harvested. The 6× His-NOB1 fusion protein was purified and analyzed by SDS-PAGE.
Mice and immunization
At 2-week intervals, a total of six adult female BALB/c mice (FMMU Animal Center, Xi'an, China), 8–10 weeks of age, were injected intraperitoneally with 400 μL of a 1:1 emulsion of NOB1 fusion protein (20 μg per mouse) and complete Freund's adjuvant (Gibco-BRL). After four booster shots, the immunity was assessed by indirect enzyme-linked immunosorbent assay (ELISA). The mice were again boosted 3 days before the fusion.
Hybridoma preparation
The fusion to SP2/0 myeloma cells was carried out as described previously.(8) The fused cells were resuspended in 40 mL of selection medium, and the aliquots of the cell suspension (100 μL) were dispensed into the wells of 96-well plates. The medium was replaced with fresh selection medium every 2 or 3 days. Finally, the hybridoma cells of positive wells were cloned by limiting dilution in aminopterin-free selection medium.
ELISAs
Ninety-six-well polystyrene plates were incubated with 20 μg/well of either 6× His-NOB1 fusion protein or control protein in phosphate-buffered saline [pH 8.0] overnight at 4°C. The plates were washed, and the unbound sites were incubated with 2% bovine serum albumin. After triplicate washing with buffer A, the wells were incubated with horseradish peroxidase (HRP)-conjugated anti-mouse IgG antibody (0.2 μg/mL) (1:1000 v/v; Sigma). The light absorbance of each well was determined at 490 nm.
Western blotting
Western blotting was carried out as described previously.(8) Briefly, a total of 60 μg of lysates were electrophoresed on 12% SDS-PAGE. The membrane was reacted with Mab and the HRP-conjugated goat anti-mouse IgG antibody. Enhanced chemiluminescence (Amersham) was added and monitored for the development of color.
Statistical analysis
All data were analyzed by χ2 test using the SPSS software package (SPSS, Inc.). A value of p < 0.05 was considered significant.
Results
Expression and purification of fusion protein
The result of restriction endonuclease analysis indicated that all the white clones were correct recombinant clones. DNA sequencing of the recombinant plasmid confirmed that the sequence of the inserted fragment was identical to the reported sequence. After being induced by IPTG, the expression of 6× His-NOB1 fusion protein was verified by SDS-PAGE and Coomassie Brilliant Blue staining.
Preparation of NOB1-specific MAb
The splenocytes from a mouse with a high-titered immune response to the fusion protein were fused with SP2/0 myeloma cells, and the resulting hybridomas were screened for the secretion of anti-NOB1-specific antibodies by ELISA. The purified recombinant 6× His-ZNRD1 fusion protein was used as a negative control in the first round of ELISA screening. As a result, hybridoma supernatants from 19 wells (19/384; 4.95%) were found to be specific to the NOB1 protein without recognizing 6× His-ZNRD1 fusion protein. Finally, one hybridoma clone was obtained and named L6 (IgG1).
Identification of MAb L6
To test the specificity of MAb L6, western blot analyses of 6× His-NOB1 fusion protein in SGC7901 cells were performed using MAb supernatant as the primary antibody. Mouse polyclonal antibodies reacting with the NOB1 fusion protein were used as a positive control, and 6× His-ZNRD1 fusion protein was used as a negative control. The results (Fig. 1) clearly showed that L6 could recognize the recombinant NOB1 protein.

Western blot analyses of NOB1 protein in SGC7901 cells using MAb L6. Lane 1: positive control (the mouse polyclonal antibodies reacting with the NOB1 fusion protein); lane 2: MAb L6; and lane 3: negative control (6× His-ZNRD1 fusion protein). MAb, monoclonal antibody.
Expression of NOB1 in different cells
The expression of NOB1 protein was detected in GES, MKN28, SGC7901, MKN45, SGC7901/VCR, and SGC7901/ADR cells (Fig. 2). β-Actin was used as a control. NOB1 protein was found upregulated in gastric cancer cells compared with normal gastric epithelial cells. NOB1 was also found upregulated in multidrug-resistant gastric cancer cells SGC7901/VCR and SGC7901/ADR compared with the drug-sensitive cells SGC7901.

Expression of NOB1 in different cells. β-Actin was used as control. Lane 1: GES cells; lane 2: MKN28 cells; lane 3: SGC7901 cells; lane 4: SGC7901/VCR cells; and lane 5: SGC7901/ADR cells.
Expression of NOB1 in gastric cancer tissues
As shown in Figure 3, the expression of NOB1 was higher in gastric cancer tissues compared with the adjacent nonneoplastic tissues.
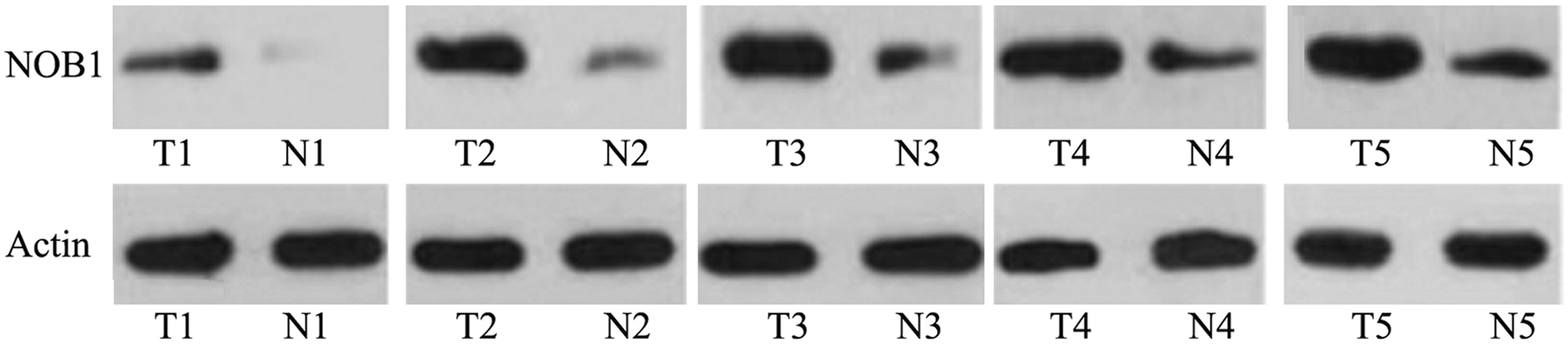
Detection of NOB1 in the gastric cancer tissues and adjacent nonneoplastic tissues by western blot. β-Actin was used as control. T1–T5: cancer tissues; N1–N5: adjacent nonneoplastic tissues. β-Actin was used as control.
Discussion
The human NOB1 gene spans about 13 kb on the genome and consists of nine exons and eight introns. It encoded a transcription-associated protein consisting of a zinc ribbon domain at its C terminus from 208 to 296aa.(9,10) The zinc ribbon domain was a functional domain required for biological activities in TFIIS and TFIIB. Since NOB1 involved a zinc ribbon domain, it was assumed to be related to transcription regulation.(11) Nob1p is required for cleavage of the 3′ end of 18S rRNA.(12) In addition, Nob1 binds the single-stranded cleavage site D at the 3′ end of 18S rRNA with its PIN domain.(13)
The expression of NOB1 was detectable in many cancers. Chen et al. have found that the positive NOB1 expression predicted poor outcome in prostate cancer.(14) Lin et al. have found that the expression of NOB1 mRNA and protein in papillary thyroid carcinoma tissue was significantly higher than that in normal thyroid tissue and benign thyroid tissue, while there was no significant difference between normal thyroid tissue and benign thyroid tumor tissue.(15) Meng et al. have found that the adenovirus-mediated siRNA targeting NOB1 inhibited tumor growth and enhanced radiosensitivity of human papillary thyroid carcinoma in vitro and in vivo.(16) Silencing NOB1 enhanced doxorubicin antitumor activity of the papillary thyroid carcinoma in vitro and in vivo. Li et al. have clarified that the aberrant expression of NOB1 in breast infiltrating ductal carcinoma was possibly involved with tumorigenesis and development, and the NOB1 protein could act as a potential biomarker for prognosis assessment of breast infiltrating ductal carcinoma.(17) Jia et al. have found that NOB1 played an essential role in renal cancer cell proliferation, and its gene expression could be a therapeutic target.(18)
In this study, we generated one effective MAb L6 against the NOB1 protein. Because the mouse were immunized with a 6× His fusion protein, it was possible that the antibodies produced by the hybridomas could be reactive to either NOB1 or 6× His. To identify hybridomas secreting MAbs against NOB1 protein, purified recombinant 6× His-ZNRD1 fusion protein was used as a negative control. L6 was proven to be valuable for detecting the denatured and native form of endogenous NOB1 protein. Using L6, NOB1 protein was first found upregulated in gastric cancer cells and tissues compared with the normal ones. Zhou et al. have found that the positive expression rate of NOB1 in gastric cancer tissue was 73%, higher than that in adjacent normal tissue (47%).(19) The result is consistent with our data. NOB1 was also first found upregulated in multidrug-resistant gastric cancer cells compared with the drug-sensitive cells. Further work should be carried out to investigate the role of NOB1 in carcinogenesis and multidrug resistance of gastric cancer.
Footnotes
Acknowledgment
This work was supported by the National Natural Science Foundation of China (no. 81171923).
Author Disclosure Statement
No competing financial interests exist.