
Abstract
Podoplanin (PDPN) is a type I transmembrane O-glycoprotein, which is known as a specific lymphatic marker. PDPN activates platelet aggregation by binding to C-type lectin-like receptor-2 (CLEC-2) on platelet. PDPN is also expressed in several normal tissues, including podocytes and type I alveolar cells. Although many monoclonal antibodies (MAbs) against human PDPN (hPDPN), mouse PDPN (mPDPN), and rat PDPN (rPDPN) have been established, useful antibodies against rabbit PDPN (rabPDPN) have not been developed. In this study, we immunized mice with the recombinant proteins of rabPDPN, and developed a novel anti-rabPDPN MAb, named PMab-32. PMab-32 could detect endogenous and exogenous rabPDPN in flow cytometry and Western blot analysis. The KD of PMab-32 was determined to be 6.2 × 10−8 M by flow cytometry. Immunohistochemical analysis showed that PMab-32 is useful for detecting podocytes, type I alveolar cells, and lymphatic endothelial cells in normal rabbit tissues. PMab-32 is expected to be useful for various rabbit experiments.
Introduction
P
Expression of PDPN has been reported in many malignant tumors such as malignant brain tumors,(14–17) oral squamous cell carcinomas,(18) lung cancers,(19) esophageal cancers,(20) malignant mesotheliomas,(21,22) testicular tumors,(23) osteosarcomas,(5,24,25) and chondrosarcomas.(5) PDPN is also associated with tumor invasion and metastasis.(26) Moreover, PDPN expression in cancer-associated fibroblasts (CAFs) is involved in poor prognosis of several malignant tumors.(27)
In this study, we immunized mice with the recombinant proteins of rabbit PDPN (rabPDPN) and developed a novel anti-rabPDPN monoclonal antibody (MAb).
Materials and Methods
Cell lines
Chinese hamster ovary (CHO)-K1 and P3U1 were purchased from American Type Culture Collection (ATCC, Manassas, VA). VX2 (rabbit skin squamous cell carcinoma) was obtained from Cell Resource Center for Biomedical Research Institute of Development, Aging and Cancer (Tohoku University, Sendai, Japan). CHO-K1 was previously transfected with human PDPN (hPDPN)-FLAG tag, mouse PDPN (mPDPN)-FLAG tag, or rat PDPN (rPDPN)-V5-His tag plasmids (CHO/hPDPN, CHO/mPDPN, CHO/rPDPN) using Lipofectamine LTX (Thermo Fisher Scientific, Waltham, MA). CHO-K1 cells were transfected with the rabbit PDPN (rabPDPN)-PA tag plasmid using a Gene Pulser Xcell electroporation system (Bio-Rad Laboratories, Philadelphia, PA). CHO-K1, CHO/rabPDPN, CHO/hPDPN, CHO/mPDPN, CHO/rPDPN, VX2, and P3U1 were cultured in RPMI 1640 medium including
Animals
Female BALB/c mice (4 weeks old) were purchased from CLEA Japan (Tokyo, Japan). Animals were housed under pathogen-free conditions. The Animal Care and Use Committee of Tohoku University approved the animal experiments described herein.
Expression and purification of recombinant rabPDPN
The rabPDPN cDNA (GenBank accession no. XM_008275367) was obtained by polymerase chain reaction (PCR) using total RNA extracted from a normal rabbit lung. The amplified cDNAs were subcloned into a pCR4-TOPO vector (Thermo Fisher Scientific). Expression construct for rabPDPN was cloned into XhoI-NotI site of pCAG-Ble PA tag-N vector (Wako Pure Chemical Industries, Osaka, Japan). This construct was verified by DNA sequencing. For the purification of rabPDPN from cell membrane, CHO/rabPDPN was plated in 100 10-cm culture dishes. Cells were detached from the plates using phosphate buffered saline (PBS) containing 1 mM EDTA. The pelleted cells were washed using PBS and solubilized by adding 10 mL of PBS containing 1% (w/v) Triton X-100 and protease inhibitor cocktail (Wako Pure Chemical Industries). The solubilized cell lysates were incubated at 4°C for 30 min and further centrifuged 15 min at 13,000 g at 4°C. The pelleted cells were solubilized by 5 mL of the same solution and the same process was repeated twice. The cleared lysates were mixed with NZ-1-Sepharose (3 mL bed volume) and incubated at 4°C for 2 h under gentle agitation.(28) The beads were then transferred to a column and washed with 60 mL of PBS. The bound protein was eluted with PBS containing 0.1mg/mL epitope peptide (EGGVAMPGAEDDVV). The elution was conducted at room temperature in a step-wise manner (3 mL ×10), where dissociation time of 5 min was given for each elution step.
Hybridoma production
BALB/c mice were immunized by intraperitoneal (i.p.) injection of 100 μg of recombinant rabPDPN together with Imject Alum (Thermo Fisher Scientific). After several additional immunizations of 50 μg, a booster injection of 50 μg was given i.p. 2 days before spleen cells were harvested. The spleen cells were fused with P3U1 cells using GenomONE-CF (Ishihara Sangyo Kaisha, Osaka, Japan). The hybridomas were grown in RPMI medium with hypoxanthine, aminopterin, and thymidine selection medium supplement (Thermo Fisher Scientific). The culture supernatants were screened using enzyme-linked immunosorbent assay (ELISA) for binding to the recombinant proteins of rabPDPN.
ELISA
Recombinant proteins of rabPDPN were immobilized on Nunc Maxisorp 96-well immunoplates (Thermo Fisher Scientific) at 1 μg/mL for 30 min. After blocking with 1% BSA/PBS, the plates were incubated with culture supernatant followed by 1:3000 diluted peroxidase-conjugated anti-mouse IgG (Dako, Agilent Technologies, Santa Clara, CA). The enzymatic reaction was conducted with a 1-Step Ultra TMB-ELISA (Thermo Fisher Scientific). The optical density was measured at 655 nm using an iMark microplate reader (Bio-Rad Laboratories).
Western blot analysis
Cell lysates (10 μg) were boiled in SDS sample buffer (Nacalai Tesque). The proteins were electrophoresed on 5–20% polyacrylamide gels (Wako Pure Chemical Industries) and were transferred onto a PVDF membrane (EMD Millipore, Billerica, MA). After blocking with 4% skim milk (Nacalai Tesque), the membrane was incubated with PMab-32, LpMab-7, PMab-1, PMab-2, and anti-β-actin (clone AC-15, Sigma-Aldrich, St. Louis, MO) and then with peroxidase-conjugated secondary antibodies (1:1000 diluted; Dako, Agilent Technologies), and developed with the ImmunoStar LD Chemiluminescence Reagent (Wako Pure Chemical Industries) using a Sayaca-Imager (DRC Co., Tokyo, Japan).
Flow cytometry
Cells were harvested by brief exposure to 0.25% Trypsin/1 mM EDTA (Nacalai Tesque). After washing with 0.1% BSA/PBS, the cells were treated with primary MAbs (1 μg/mL) for 30 min at 4°C followed by treatment with Oregon green-conjugated anti-mouse IgG or anti-rat IgG (1:1000 diluted; Thermo Fisher Scientific). Fluorescence data were collected using a Cell Analyzer EC800 (Sony, Tokyo, Japan).
Determination of binding affinity by flow cytometry
Binding affinity was determined as described previously.(17,29) Briefly, cells (2 × 105 cells) were resuspended at 100 μL of serially diluted antibodies (600 pg/mL-10 ug/mL), followed by secondary antibodies (Thermo Fisher Scientific). Fluorescence data were collected using a Cell Analyzer EC800. The dissociation constants (KD) were obtained by fitting the binding isotherms using the built-in one-site binding models in GraphPad PRISM 6 (GraphPad Software, La Jolla, CA).
Determination of binding affinity by surface plasmon resonance
To determine the affinity, biotinylated recombinant rabPDPN was immobilized on the SA sensor chip for analysis using the BIAcore X100 system (GE Healthcare, Piscataway, NJ). The running buffer was PBS containing 0.005% Tween-20 (PBST). The PMab-32 was diluted in PBST (6.25, 12.5, 25, 50, 100 nM), and was passed over the biosensor chip. The binding was monitored by the single-cycle kinetics method. The binding curves were analyzed by nonlinear curve-fitting using the bivalent analyte model of the BIAcore X100 Evaluation Software (BIAcore). The affinity constant (KA1) at equilibrium was calculated as KA1 = kassoc1/kdiss1, and the dissociate constant (KD1) was determined as 1/KA1.
Immunohistochemical analyses
Four-μm-thick histologic sections were deparaffinized in xylene and rehydrated. Then, they were autoclaved in citrate buffer (pH 6.0; Dako, Agilent Technologies) for 20 min. Sections were incubated with 1 μg/mL of PMab-32 for 1 h at room temperature, followed by treatment with Envision+ kit for 30 min (Dako, Agilent Technologies). Color was developed using 3, 3-diaminobenzidine tetrahydrochloride (DAB; Dako, Agilent Technologies) for 1 min, and then the sections were counterstained with hematoxylin (Wako Pure Chemical Industries).
Results
Production of monoclonal antibody against rabbit podoplanin
We first immunized mice with the recombinant protein of CHO/rabPDPN, and the ELISA screening was performed. Among 127 ELISA-positive wells, 55 wells reacted with CHO/rabPDPN in flow cytometry. Twenty-four of these 55 wells reacted with VX2 rabbit skin squamous cell carcinoma cell line, which expresses the endogenous rabPDPN in flow cytometry. Seventeen of 24 wells of IgG1 subclass were cloned by limiting dilution. Then, PMab-32 (mouse IgG1, kappa) was established. PMab-32 reacted with CHO/rabPDPN, but not with CHO/hPDPN, CHO/mPDPN, CHO/rPDPN, and CHO cells in flow cytometry (Fig. 1). LpMab-7,(24,30,31) PMab-1,(32,33) and PMab-2(34) specifically reacted with CHO/hPDPN, CHO/mPDPN, and CHO/rPDPN, respectively. PMab-32 also recognized endogenous rabPDPN, which is expressed in VX2.
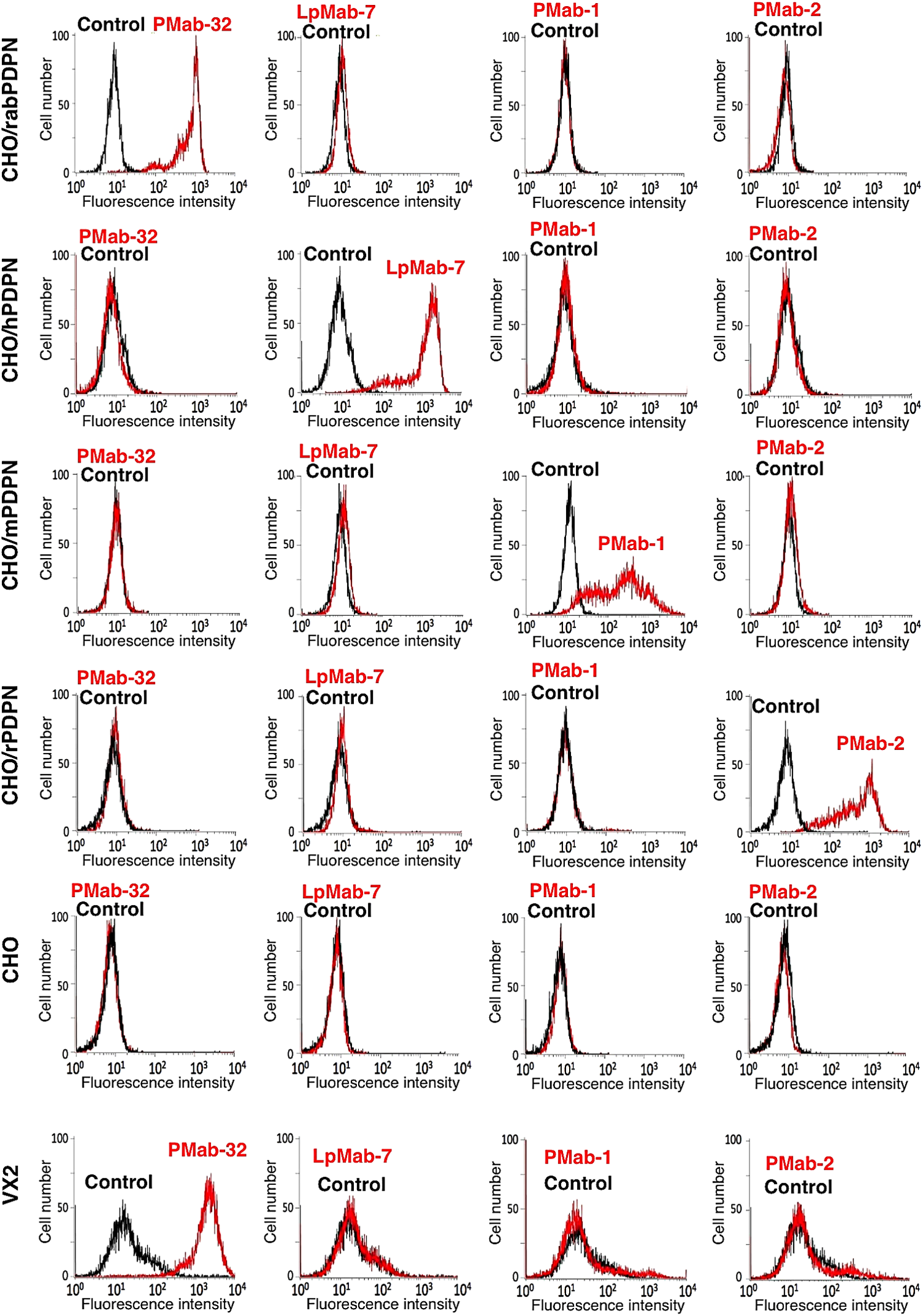
Flow cytometric analysis by anti-PDPN MAbs. CHO/rabPDPN, CHO/hPDPN, CHO/mPDPN, CHO/rPDPN, CHO, and VX2 were treated with PMab-32, LpMab-7, PMab-1, and PMab-2, followed by treatment with Oregon green-conjugated anti-mouse IgG or anti-rat IgG. Fluorescence data were collected using a Cell Analyzer EC800. Red line, PMab-32, LpMab-7, PMab-1, or PMab-2; black line, negative control.
PMab-32 detected about a 40 kDa band of CHO/rabPDPN, but not with CHO/hPDPN, CHO/mPDPN, CHO/rPDPN, and CHO cells in Western blot analysis (Fig. 2). In contrast, LpMab-7, PMab-1, and PMab-2 detected about 40 kDa bands of CHO/hPDPN, CHO/mPDPN, and CHO/rPDPN, respectively. PMab-32 also detected endogenous rabPDPN, which is expressed in VX2 rabbit skin squamous cell carcinoma cell line, indicating that PMab-32 is also useful in Western blot analyses to detect rabPDPN. The molecular weight of endogenous rabPDPN is smaller than that of PA-tagged rabPDPN.
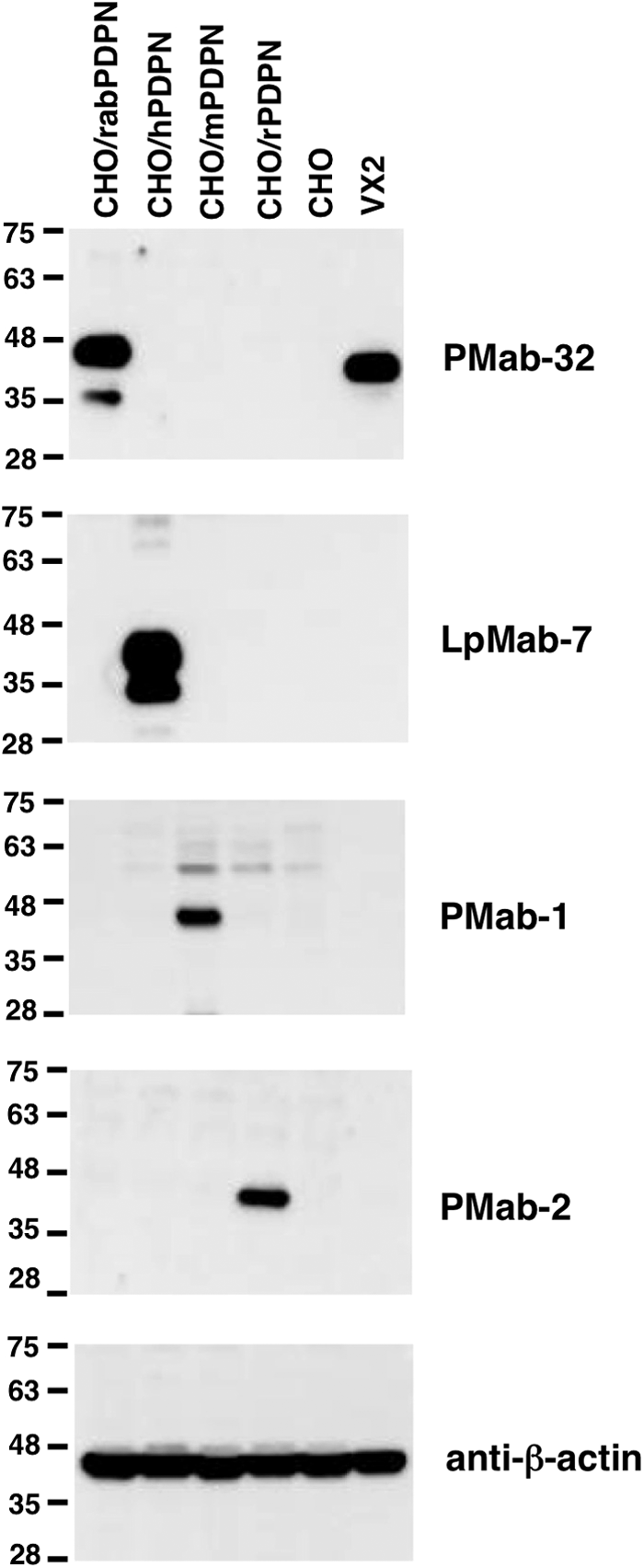
Western blot analysis by anti-PDPN MAbs. Cell lysates (10 μg) were electrophoresed on 5–20% polyacrylamide gels and transferred onto a PVDF membrane. After blocking, the membrane was incubated with 1 μg/mL of MAbs (PMab-32, LpMab-7, PMab-1, PMab-2, and anti-β-actin), and then with peroxidase-conjugated secondary antibodies; the membrane was developed with ImmunoStar LD Chemiluminescence Reagent and detected using a Sayaca-Imager.
We further performed a kinetic analysis of the interaction of PMab-32 with CHO/rabPDPN using flow cytometry. As shown in Figure 3A, KD of PMab-32 was determined to be 6.2 × 10−8 M using flow cytometry against CHO/rabPDPN. BIAcore analysis showed that KD1 of PMab-32 was determined to be 1.1 × 10−7 M (Fig. 3B).

Determination of binding affinity using flow cytometry and surface plasmon resonance. (
Immunohistochemical analysis using PMab-32
We investigated whether PMab-32 can detect rabPDPN of normal tissues such as kidney, lung, and small intestine. As shown in Figure 4, PMab-32 reacted with podocytes of kidney, type I alveolar cells of lung, and lymphatic endothelial cells of small intestine. Lymphatic endothelial cells of other tissues were also stained by PMab-32 (data not shown). The antigen retrieval procedure is essential for staining of type I alveolar cells of lung and lymphatic endothelial cells, whereas podocytes were stained by PMab-32 without antigen retrieval (data not shown). These data show that PMab-32 is useful for immunohistochemistry using paraffin-embedded tissues; however, antigen retrieval is necessary for using a lymphatic endothelial cell marker.
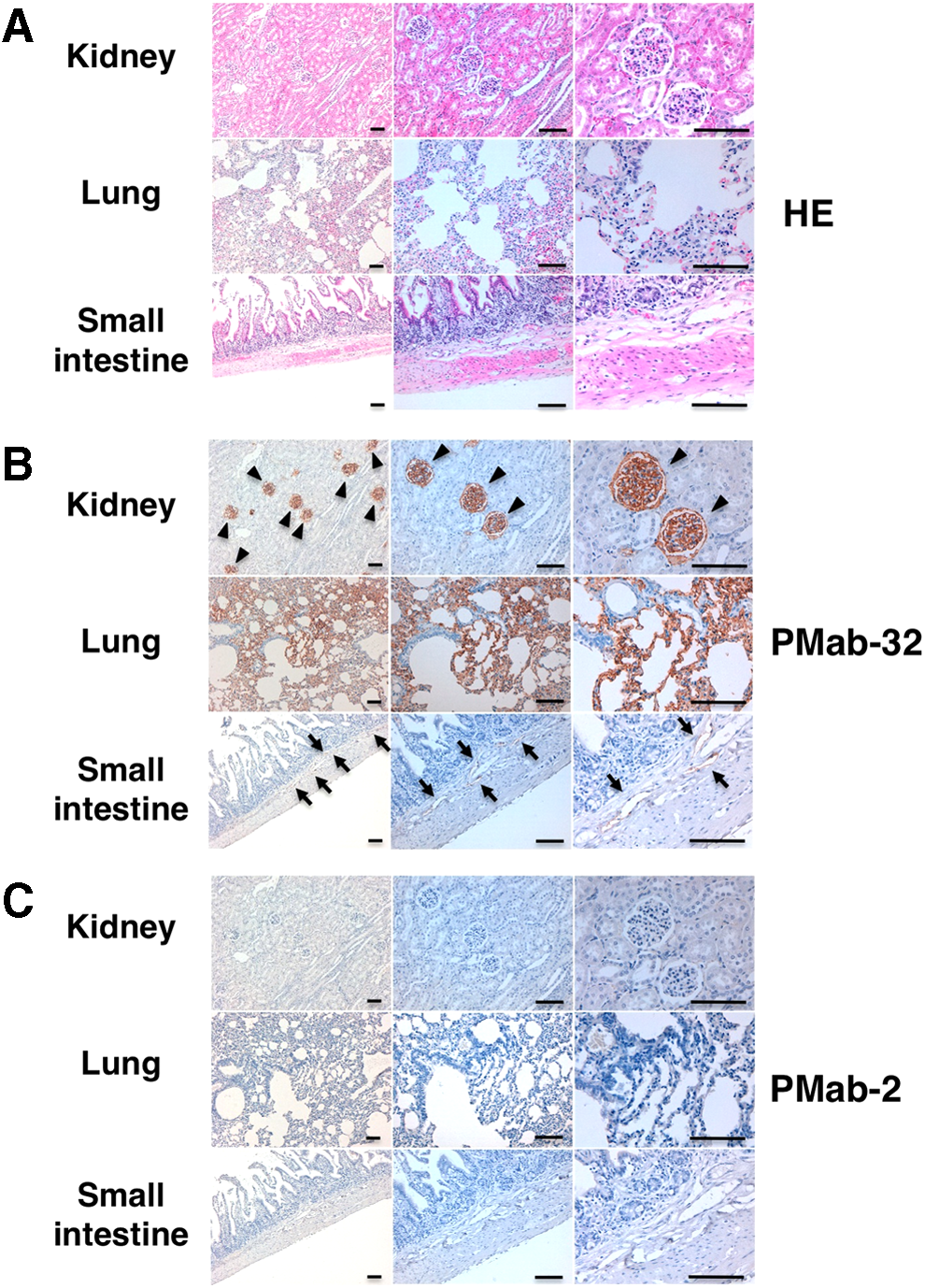
Immunohistochemical analysis by PMab-32. (
Discussion
PDPN is expressed in human fetal rib and chondrocytes of the proliferative and hypertrophic regions of the growth plate.(35) Binding of the cytoplasmic tail of PDPN to the ERM proteins may change cytoskeletal organization, which alters the phenotype of PDPN expression cells. This may contribute to morphological changes in the rudiment cartilages, which leads to the establishment of primary ossification centers during the endochondral ossification.
Rabbits are one of the most commonly used animals in orthopedic research such as osteoarthritis, osteochondritis dissecans, bone formation, bone healing, and cartilage repair.(36–40) Several specific MAbs against osteonectin, collagen types I and II, and osteocalcin were used in those studies. Although many MAbs against hPDPN,(17,24,31,41–46) mPDPN,(32) and rPDPN(34) have been established, useful antibodies against rabbit PDPN, especially in immunohistochemical analysis, have not been developed. Many clones, which can be used only in Western blot and flow cytometry, but not in immunohistochemistry, have been produced before PMab-32 was established (data not shown). Because we established specific MAbs against hPDPN, mPDPN, and rPDPN by immunizing mice or rats with synthetic peptides of PLAG domains, we applied the same method in several trials; synthetic peptides including the PLAG domain of rabPDPN were used for production of specific MAbs. However, established 24 clones (PMab-8 through PMab-31) showed strong background staining in immunohistochemistry (data not shown). Twenty of 24 clones (83%) were determined to be IgG class; therefore, amounts and time of immunization were enough for affinity maturation. Then, we changed the methods in subsequent experiments. Firstly, purified membranous rabPDPN from CHO/rabPDPN cells was used for immunization. Secondly, we used an endogenous rabPDPN-expressing cell line VX2 for second screening of flow cytometry. The binding affinity of PMab-1, PMab-2, and NZ-1, which was determined using flow cytometry, was 3.2 × 10−9 M, 5.9 × 10−10 M, and 2.9 × 10−9 M, respectively.(34) Although the binding affinity of PMab-32 (6.2 × 10−8 M using flow cytometry) was lower than PMab-1, PMab-2, and NZ-1; PMab-32 is highly sensitive in Western blot, flow cytometry, and immunohistochemistry. Taken together, PMab-32 reacts with rabPDPN-expressing normal cells; therefore, it could be useful for uncovering the PDPN function in the rabbit model.
Footnotes
Author Disclosure Statement
The authors have no financial interests to disclose.
Acknowledgments
We thank Noriko Saidoh, Hazuki Kanno, and Kanae Yoshida for their excellent technical assistance. We also thank Satoru Konnai, Masahiro Maruyama, and Akiko Kunita for preparing paraffin block of rabbit tissues. This work was supported in part by the Platform for Drug Discovery, Informatics, and Structural Life Science (PDIS) from Japan Agency for Medical Research and development, AMED (Y.K.); the Basic Science and Platform Technology Program for Innovative Biological Medicine from AMED (Y.K.); the Regional Innovation Strategy Support Program from the Ministry of Education, Culture, Sports, Science and Technology (MEXT) of Japan (Y.K.); and JSPS KAKENHI (grant nos. 26440019 [M.K.K.] and 25462242 [Y.K.]).