
Abstract
Bone marrow stromal cells (BMSCs) are very useful model systems for a better understanding of cell behavior and differential gene expression. Up to now, there have not been specific markers and MAbs for BMSCs that hamper the identification and isolation of BMSCs populations. In this study, chicken BMSCs were isolated from 1-day-old Beijing fatty chickens by adherent culture. After biological characteristics were detected, the chicken BMSCs were used to immunize BALB/c mice to prepare BMSCs-specific monoclonal antibodies (MAbs) by the routine hybridoma technique. These MAbs were characterized by FACS analysis, immunocytochemistry, immunohistochemistry, subtype identification, and Western blotting assay and were used to explore markers of chicken BMSCs. Our data showed that BMSCs expressing antigens CD29, CD44, and CD105, but not expressing antigens CD34, CD45, and CD11b, could be isolated from postnatal chicken bone marrow and hold great potential for multiline age differentiation. Meanwhile, we obtained two hybridoma cell lines secreting chicken BMSCs-specific MAbs (named CHK1 and CHK2), which specifically recognized the surface antigens expressed on chicken BMSCs. According to our subtype identification, heavy chains of CHK1 and CHK2 were typed as IgG1 and IgG2b, respectively; all the light strands were kappa subtype. MAbs CHK1 and CHK2 can be used to develop the detection assay and to discover novel biomarkers of chicken BMSCs.
Introduction
A
The adherent cells from the bone marrow that are called bone marrow stromal cells (BMSCs) have been used for bone tissue engineering, but this population is heterogeneous and only a limited number of cells (putative stem/progenitor cells) can form a colony (osteogenic). Only a small proportion of those adherent cells showed multipotency, which has been repeatedly confirmed at the single cell-derived colony level, and the other cells stay in a range of differentiation status.(2–4) Accordingly, we prefer to call the cells “BMSCs.”
Since the 1990s, researchers have attempted to explore specific markers for MSCs and prepared some MAbs, such as SH2, SH3, SH4, and STRO-1. Unfortunately, none of these markers is really specific for MSCs.(5,6) To date, the minimal criterion that has been proposed to define MSCs is that the cells are positive for CD73, CD90, and CD105 and negative for CD14, CD19, CD34, CD45, and HLA-DR.(7–9) So, the lack of common standards and a precise definition of MSCs are major obstacles in the research and application of MSCs. Thus, the identification of a definitive marker is of the utmost importance for the prospective isolation of MSCs.
Chicken is the first farm animal with a completely sequenced genome. Because of embryonic development in ovo rather than in utero, chicken is an important model for studies of embryology, immunology, oncology, cell biology, virology, and gene regulation. BMSCs are also used for gene transfer, producing medical antibodies. In addition, owing to their easy obtainability and suitability for large-scale preparation, chicken BMSCs were chosen as our experimental material.
There is a requirement for single, specific markers of the unequivocal immunophenotyping of BMSCs. In this study, our aim was to produce MAbs as a specific probe to identify new surface molecules of BMSCs. The BMSCs were isolated from hatched, specific pathogen-free chickens and characterized. Two chicken BMSCs-specific MAbs, namely CHK1 and CHK2, were produced by the mice immunized with chicken BMSCs, and they were used to detect BMSCs by IHC and Western blotting assay. We expected that these antibodies could be used for the development of a detection assay and to discover novel biomarkers of chicken BMSCs, and that they will serve as an important tool for BMSCs research.
Materials and Methods
Experimental animals
Six-week-old female BALB/c mice were purchased from the Laboratory Animal Center, Chinese Academy of Sciences, Shanghai, China. Beijing fatty chickens were obtained from the State Key Laboratory of Bioreactor Engineering, School of Biotechnology, East China University of Science and Technology, Shanghai, China. The mice for the activity test and immunization were kept under approved standard conditions for experimental animals. All experimental procedures involving animals were conducted in accordance with the protocols and guidelines for agricultural animal research imposed by the Committee for Ethics of China.
Cell lines and tissue culture
MSC (human, mouse, and rabbit) cell lines, commercially available, were obtained from ATCC (Rockfeller, MD); MSC cells were cultured in L-DMEM with 10% fetal bovine serum (FBS). All cell lines were grown at 37°C in a 5% CO2 incubator.
Chicken umbilical cord mesenchymal stem cells (UC-MSCs) were prepared from 16-day-old chicken embryos. Fetal abdominal wall umbilical and chorioallantoic membrane cord-like structures were dissociated in 0.25% trypsin, 0.02% EDTA at room temperature (RT) for 30 minutes. The cell suspension was passed through a sieve to remove cell aggregates. Cells were collected and seeded at 1.0 × 106 cells/cm2 in T-25 flasks with 5 mL L-DMEM from GIBCO Co. (Grand Island, NY). Cultures were maintained at 37.5°C in a humidified atmosphere containing 95% air and 5% CO2.
Chicken non-MSCs (muscle cells, adipocytes, osteoblasts, cardiocytes, and endotheliocytes) were prepared from 1-day-old chick tissue (muscle tissue, groin fat pads, skull with periosteum, heart tissue, and umbilical vein) with 0.25% trypsin, 0.02% EDTA with Collagenase, I (0.1%) at RT. The cell suspension was passed through a sieve to remove cell aggregates. Cells were collected and seeded at 1.0 × 106 cells/cm2 in T-25 flasks with 5 mL DMEM from GIBCO Co. Cultures were maintained at 37.5°C in a humidified atmosphere containing 95% air and 5% CO2.
Culture and biological assay of chicken BMSCs
Bone marrow samples were drained from the tibia of a 1-day-old chicken. First, the marrow was broken up by repeatedly pipetting, and the cell suspension was passed through a sieve to remove cell aggregates. Cells were pelleted by centrifugation, then resuspended with phosphate-buffered saline (PBS), and fractionated on a density gradient Ficoll-Hypaque (1.077 g/mL, 100%) from GE Healthcare Co. (Fairfield, CT) for 20 minutes at 400 g. The interface was collected and seeded at 1.0 × 106 cells/cm2 in T-25 flasks with 5 mL L-DMEM. Cultures were maintained at 37.5°C in a humidified atmosphere containing 95% air and 5% CO2. The nonadherent cells were removed after 3 days, and the culture medium was changed every 3 days. When cultures became 80% confluent, cells were detached with 0.25% trypsin and 0.02% EDTA (1:1) for 3–5 minutes at 37.5°C, and they were reseeded at 8.0 × 103 cells/cm2 in T-25 flasks with 5 mL L-DMEM for serial passaging.
BMSCs of passages 2 were harvested and made into cell suspensions; according to the method of Li,(10) 2.5 × 104 cells/mL were seeded onto 24-well plates. After culturing for 7 days, three wells were monitored daily for cell concentration until a plateau phase was reached. The cell growth curve of BMSCs was plotted.
The different surface antigens of MSCs were detected by immunofluorescence staining. MSCs of passages 3, 6, and 8 BMSCs were fixed in 4% paraformaldehyde/PBS for 15–20 minutes, then blocked for 10 minutes with methanol containing 0.1% Triton X-100 and 0.3% H2O2 to eliminate endogenetic peroxides, and incubated in a working solution of goat serum for 15 minutes to block nonspecific binding. Then, the BMSCs were incubated with primary antibodies such as CD29, CD34, CD45, and CD105 from Peprotech, Inc. (Rock Hill, NJ) at 4°C overnight; they were incubated with secondary antibodies that were conjugated with FITC from Chemicon International, Inc. (Temecula, CA) and with 0.01 mol/L PBS as a negative control, BMSCs were stained directly as a blank control. Fluorescence images were observed by a fluorescence microscope.
Immunophenotypic analysis of chicken BMSCs was performed by FACSCalibur from Becton Dickinson (San Jose, CA) with CELLQuest Pro software from Becton Dickinson. Cells were stained with the FITC-labeled anti-human CD29, CD105, and CD106 monoclonal fluorescent antibodies from Peprotech, Inc. to determine the percentage of positive cells within the sample. Gating was performed according to size and granulation using forward-scatter versus side-scatter (FSC/SSC) dot plots.
Differentiation assays of chicken BMSCs
Osteogenic differentiation was assayed according to the method of Khatri.(11) BMSCs in the induction group were incubated in DMEM media with inducers (5.5 × 10−2 mM β-mercaptoethanol, 10 mM β-sodium glycerophosphate, 10−8 mM dexamethasone, 50 mg/L vitamin C) and 10% FBS. BMSCs in the control group were cultured in expansion media. Alkaline phosphatase (ALP) levels were measured by the Gomori Ca-Co method after 2 weeks, and calcium nodus were determined by Alizarin Red staining after 3 weeks and further determined by Von Kossa silver staining.
Adipogenic differentiation was assayed according to the method of Jaiswal.(12) Adipogenic differentiation was induced by culturing confluent BMSCs cultures in DMEM supplemented with 10% FBS, 1 μM dexamethasone, 10 μg/mL insulin, and 0.5 mM IBMX for 21 days. The medium was completely replaced every 3–4 days.
Adipogenic potential was assessed by Oil Red O staining and immunofluorescence staining. The immunofluorescence staining was the same as that mentioned earlier. The primary antibodies were the anti-PPARγ antibodies from Chemicon International, Inc., and the secondary antibodies were conjugated with Cy5 from Chemicon International, Inc.
Immunization and hybridoma production
Generation of MAbs was assayed according to the method of Sepehr.(13) For the initial immunization, BMSCs were harvested and resuspended in physiological saline. The suspension containing 1.0 × 106 BMSCs/mL was emulsified with Complete Freund's Adjuvant (CFA) in a 1:1 (v/v) ratio, and it was then subcutaneously injected at five points on the mouse. Subsequent boosts were done using BMSCs suspensions emulsified in Incomplete Freud's Adjuvant (IFA) instead of CFA once a week. The serum was collected from the immunized mice on the seventh day after the fourth immunization, and it was tested for the value of antibodies by immunocytochemical staining of BMSCs. The fifth immunization was boosted with double the amount of BMSCs after the fourth immunization. On the fourth day after the fifth injection, the spleen cells were isolated to produce hybridomas.(14)
SP2/0 cells (laboratory preservation) were grown in DMEM media containing 20% FBS from GIBCO Co. The splenocytes and the myeloma cells were mixed by tapping the tube and washed twice with PBS; then, 1 mL DMEM media containing 50% (v/v) PEG 2000 was added with gentle shaking over 1 minute; next, 1 mL DMEM media was slowly added to dilute the mixture just mentioned; then, 15 mL DMEM media was added slowly to stop the fusion reaction; and finally, the mixture was centrifuged for 10 minutes at 800 rpm. The fused cell pellet was resuspended in 100 μM hypoxantine, 0.4 μM aminopterin, and 16 μM thymidine from Sigma-Aldrich, Inc. (Shanghai, China) medium and it was distributed (100 μL/well) into the 96-well tissue culture plates.
Hybridoma selection
Chicken BMSCs that were cultured in 96-well culture plates for 3 days were fixed with 4% paraformaldehyde, blocked with 3% goat serum, and finally stained with hybridoma supernatants that were collected from the hybridoma cultures and an FITC-conjugated goat anti-mouse secondary antibody from Chemicon International, Inc. Positive clones were subcloned by limiting the dilution in 96-well plates. Two weeks later, the remaining positive clones were propagated, and their culture supernatants were collected.
Specific immunoreaction detection of MAbs
A specific immunoreaction of MAbs was detected by immunofluorescence staining according to the method of Li et al.(15) The primary BMSCs were fixed in 4% paraformaldehyde/PBS for 15 minutes, then blocked for 10 minutes with methanol containing 0.1% Triton X-100 and 0.3% H2O2 to eliminate endogenetic peroxides, and incubated with a working solution of goat serum for 15 minutes to block nonspecific binding. Then, the cells were incubated with supernatant solution derived from two hybridoma cell lines that served as primary antibodies at 4°C overnight and secondary antibodies goat anti-mouse IgG conjugated with FITC from Peprotech, Inc. Next, 0.01 M PBS was used as a negative control in the place of primary antibodies and BMSCs were stained directly as a blank control. Antibodies produced by the two hybridoma cell lines were analyzed for cross-reactivity with heterologous MSCs (such as human, mouse, and rabbit), homologous UC-MSCs, and non-MSCs (such as muscle cells, adipocytes, osteoblasts, cardiocytes, and endotheliocytes). Fluorescence images were observed by a laser passing TE2000-U confocal microscopy from Nikon, Inc. (Tokyo, Japan).
FACS of MAbs (CHK1 and CHK2)
Chicken BMSCs and UC-MSCs, human MSCs, and mouse MSCs were prepared into cell suspensions, and primary antibody (CHK1, CHK2) staining buffer was added as a control. Cells were incubated at 4°C for 30 minutes. Two hundred microliter staining buffer wash was added twice. Supernatants were discarded, and 100 μL FITC-conjugated secondary antibody was added. The cells were incubated at 4°C for 30 minutes and then washed twice as described earlier but with PBS. One hundred microliter PBS containing 1% paraformaldehyde was added to each well to fix the cells, and it was then analyzed by flow cytometry 30 minutes later.
The chicken BMSCs were stained with primary antibody (CHK1, CHK2), and cells were incubated at 4°C for 30 minutes. The cells were washed twice as described earlier but with PBS, and then, FITC-conjugated secondary antibodies, CD34-PE and CD44-APC were added. The rest of the steps followed according to those described earlier.
IHC assay of MAbs
The paraffin sections were prepared according to the standard protocol.(16) Endogenous peroxidase was inactivated with 3% hydrogen peroxide for 10 minutes. The sections were blocked with a blocking solution (PBS supplemented with 10% goat serum and 1% BSA) for 1 hour at 37°C and incubated with primary antibodies that were derived from the supernatant solution of two hybridoma cell lines for 1 hour at 37°C in a humidified chamber. After being washed with PBS, the sections were incubated with an HRP-conjugated goat anti-mouse secondary antibody for 1 hour at 37°C, then washed, and finally incubated with a freshly prepared 3,3′-diaminobenzidine (DAB) substrate solution for 5 minutes at 37°C for visualization.
Subtype identification of MAbs
The subtypes of MAbs were detected using an SBA Clonotyping™ System/HRP kit from Southern Biotech, Inc. (Birmingham, AL) according to the manufacturer's protocol.
Analysis of reactivity and cross-reactivity of MAbs by Western blotting
Lysates of the primary BMSCs, homologous UC-MSCs, and heterologous MSCs of human and mouse were used to assay the reactivity and cross-reactivity of MAbs by Western blotting according to the method of Tan et al.(17) In brief, the primary BMSCs or homologous UC-MSCs and heterologous MSCs of human and mouse were resuspended in Cell Lysis Buffer from Sigma-Aldrich, Inc. that was complemented with protease inhibitors from Roche, Inc. (Shanghai, China). The proteins were separated on sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), and they were transferred onto PVDF membrane from GE Healthcare, Inc. (Shanghai, China) using a semi-dry electrotransferring system from Bio-Rad, Inc. (Berkeley, CA). The immunoreaction was detected with X-ray film from Kodak, Inc. (Rochester, NY) after treatment of the blots with a chemiluminescent reagent ECL-plus from Thermo Fisher Scientific, Inc. (Waltham, MA).
Results
Cultivation and biological assay of chicken BMSCs
We isolated chicken BMSCs from the bone marrow based on their characteristic property of attaching to a plastic surface. Most of the cultivated BMSCs were triangle-like or shuttle-like cells, and the population was heterogeneous. The initial adherent spindle-shaped cells appeared as individual cells on the third day of culture (Fig. 1A). Within 2 weeks, the cells reached 70%–80% confluence, and they appeared in a concentric circle radiation distribution (Fig. 1B). The growth curves of BMSCs were S shaped (Fig. 1C). BMSCs arrived in the exponential growth phase after 2 or 3 days, and they reached the platform growth phase after 6 days. These cells were continuously in culture for >3 months. These data indicated that chicken BMSCs were capable of high proliferation.
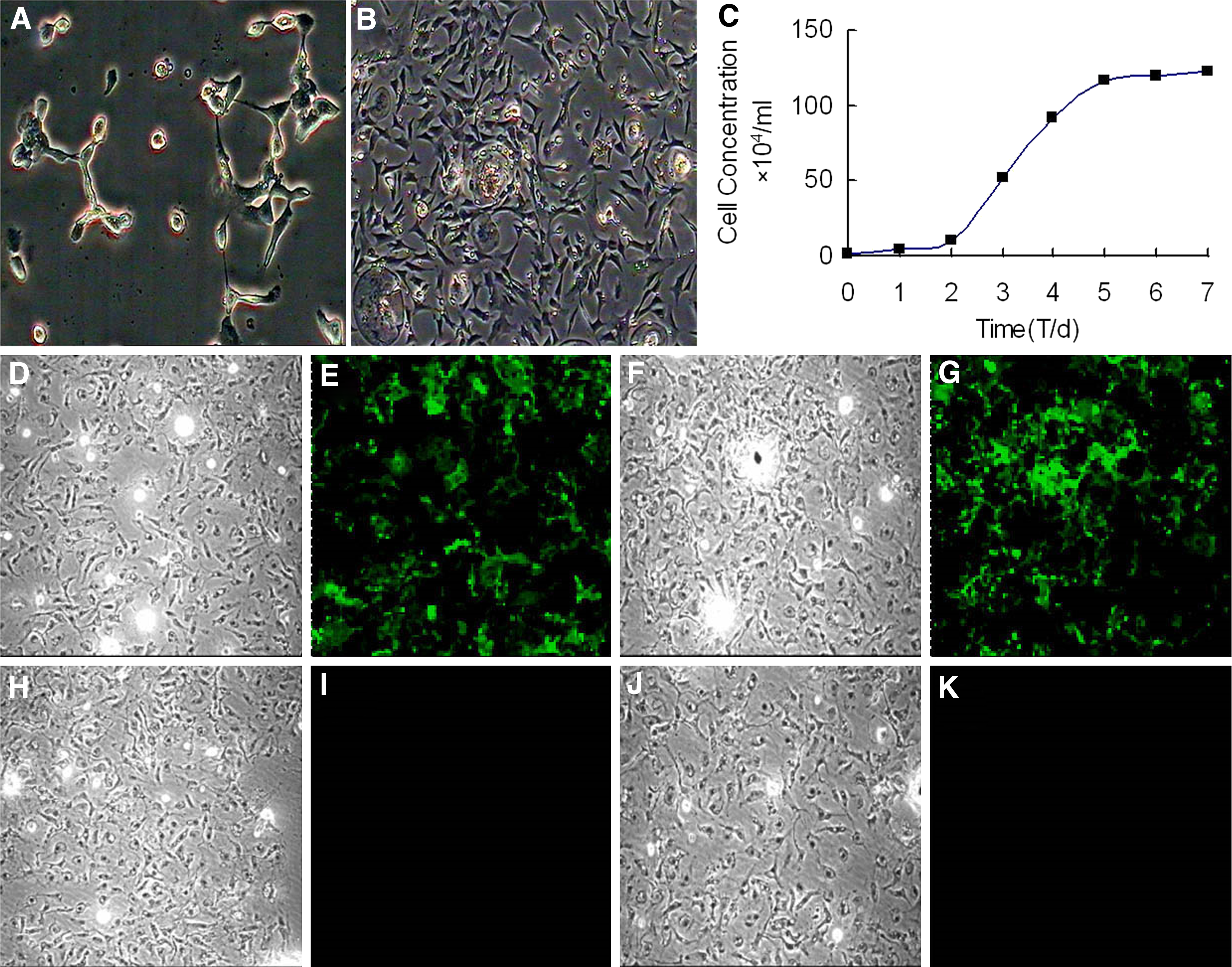
Morphology and biological assay of chicken BMSCs.
Immunofluorescence staining results showed that passage 3 BMSCs expressed antigens CD29 (Fig. 1D, E) and CD105 (Fig. 1F, G), but they did not express antigens CD34 (Fig. 1H, I) and CD45 (Fig. 1J, K), which are hematopoietic markers. There was no significant difference in the positive rates of passages 3, 6, and 8 (data not shown).
FACS results indicated that BMSCs consistently expressed stem cell markers, CD29, CD44, and CD 105, but they did not express hematopoietic markers CD34 and CD45 and monocytic marker CD11b (Fig. 2A).
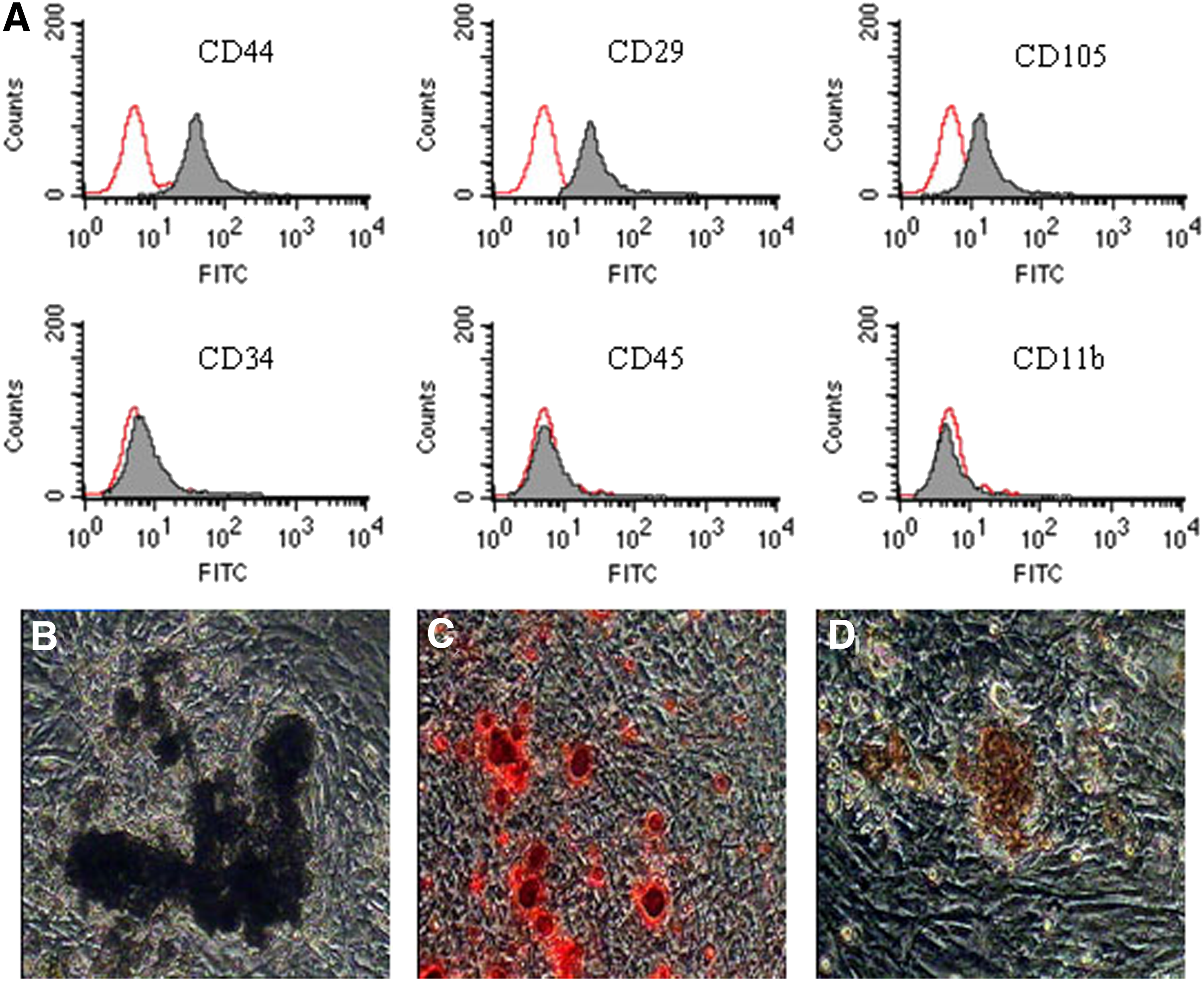
Phenotype identification and osteogenic differentiation of chicken BMSCs (100 × ).
To examine the multipotentiality of chicken BMSCs, the cells were cultured in the media that were specific for the induction of osteocytes and adipocytes. After the induction with osteoblast inducers, BMSCs showed significant changes in appearance. Some BMSCs changed from fusiform to three dimensional, becoming larger and polygon shaped from the third day after the induction. Afterward, the population of triangle or polygonal cells increased, and then, these cells grew into multi-layers; meanwhile, many crystal particles could be seen. ALP level was determined by the Gomori Ca-Co method after BMSCs had been induced for 2 weeks. Brown-black particles were found to be present in the cytoplasm (Fig. 2B). During the third week, the induced cells became bright red in the positive region after being stained with Alizarin Red (Fig. 2C). With the extension of incubation time, the number of nodules gradually increased, and Von Kossa staining was positive (Fig. 2D).
To study the generation of adipocytes, 70%–80% confluent cultures were incubated with adipocyte induction media. After being induced with induction media, the cells gradually displayed a perinuclear accumulation of lipid vacuoles, and their shape changed from long fusiform cells to round or polygonal cells (Fig. 3A). The cells became bright red in the positive region after being stained with Oil Red O (Fig. 3B, C). Meanwhile, the lipid droplets appeared; the expression of adipogenic-specific antigen PPARγ was positive (Fig. 3D, F), and that of the negative control was negative (Fig. 3G, I), thereby confirming the adipogenic differentiation of BMSCs. According to the morphology and differentiation potentials, the cells derived from the chicken marrow were identified as BMSCs.
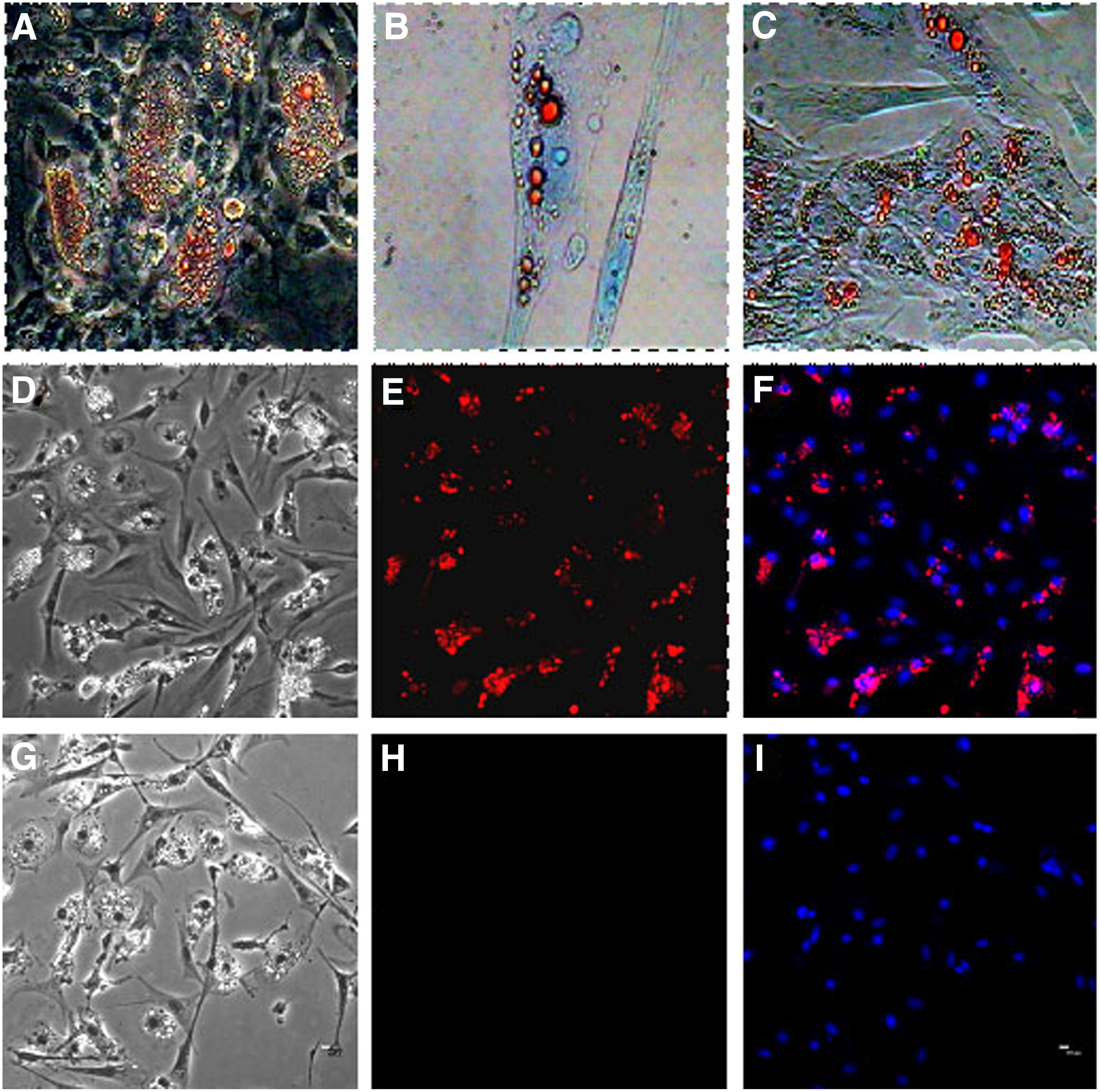
Identification of adipogenic differentiation of chicken BMSCs. Chicken BMSCs were cultured in adipogenic medium.
Isolation and selection of hybridoma cells
In this study, polyclonal hybridomas were generated in 1920 wells (96-well plates × 20 plates) and selected with immunocytochemistry (ICC) testing. Supernatants from 28 wells were positive for BMSCs. After subcloning, two hybridoma cell lines secreting chicken BMSCs-specific MAbs were finally obtained, namely CHK1 and CHK2, respectively.
Specificity of MAbs (CHK1 and CHK2)
Immunocytochemistry
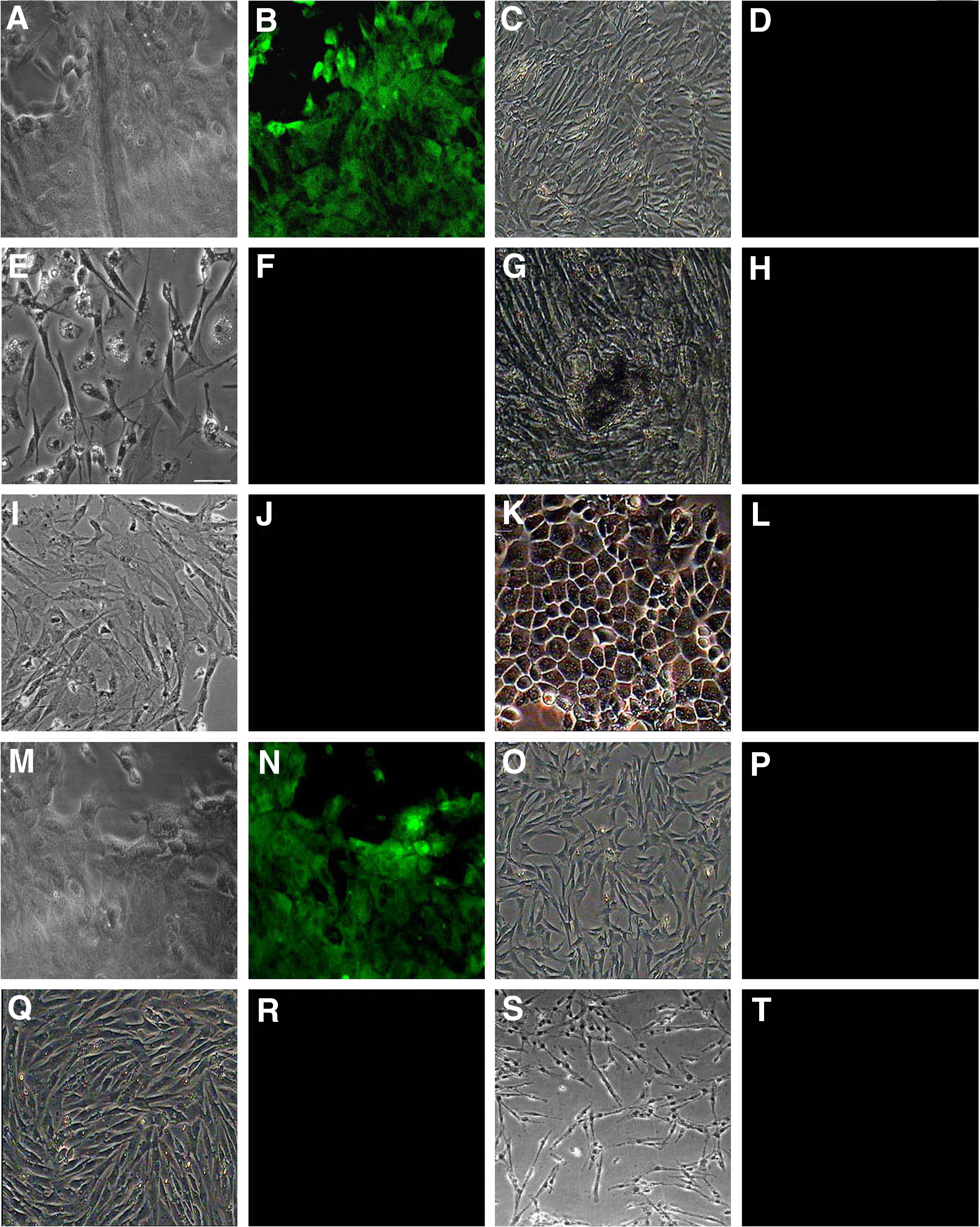
Investigations on cross-reactivity of the applied antibody are crucial for the assessment of the results. Therefore, a number of other cells, structurally related to BMSCs, were tested with an immunofluorescence assay to characterize the properties of the antibody. The CHK1 exhibited bright staining of passage 2 chicken BMSCs by immunofluorescence staining (Fig. 4A, B). The positive rate of CHK1 on chicken BMSCs was 96.35%. None was obviously reactive with non-BMSCs of muscle cells (Fig. 4C, D), adipocytes (Fig. 4E, F), osteoblasts (Fig. 4G, H), cardiocytes (Fig. 4I, J), and endotheliocytes (Fig. 4K, L). The CHK1 exhibited bright staining of chicken UC-MSCs by immunofluorescence staining (Fig. 4M, N). The positive rate of CHK1 on chicken UC-MSCs was 94.5%. None was obviously reactive with heterologous MSCs of mouse (Fig. 4O, P), human (Fig. 4Q, R), and rabbit (Fig. 4S, T).

Characterization specificity of CHK1 by immunofluorescence assay.
The CHK2 exhibited bright staining of passage 2 chicken BMSCs by immunofluorescence staining (Fig. 5A, B). The positive rate of CHK2 on chicken BMSCs was 92.7%. None was obviously reactive with muscle cells (Fig. 5C, D), adipocytes (Fig. 5E, F), osteoblasts (Fig. 5G, H), cardiocytes (Fig. 5I, J), and endotheliocytes (Fig. 5K, L). The CHK2 exhibited bright staining of chicken UC-MSCs by immunofluorescence staining (Fig. 5M, N). The positive rate of CHK2 on chicken UC-MSCs was 91.5%. None was obviously reactive with heterologous MSCs of mouse (Fig. 5O, P), human (Fig. 5Q, R), and rabbit (Fig. 5S, T).
Characterization specificity of CHK2 by immunofluorescence assay.
The data demonstrated that the CHK1 and CHK2 were highly specific for chicken BMSCs. None was obviously reactive with heterologous BMSCs (such as human, mouse, and rabbit), and non-BMSCs (such as cardiocytes, endotheliocytes, and muscle cells), but homologous UC-MSCs were obviously reactive with CHK1 and CHK2. CHK1 and CHK2, the most strikingly positive clones as judged by tests, are the subjects of this article. The two MAbs showed similar staining patterns on the BMSCs. Their targeted proteins for CHK1 and CHK2 were located on the membrane.
FACS
The specificity of two MAbs (CHK1 and CHK2) was identified by FACS. Two MAbs were found to react with chicken BMSCs and chicken UC-MSCs, and the positive populations of CHK1 and CHK2 were 95.28% and 92.78% in chicken BMSCs (Fig. 6A, B), and 93.80% and 93.61% in chicken UC-MSCs (Fig. 6C, D), respectively; however, there was no reaction with human MSCs and mouse MSCs (Fig. 6E, I). These data indicated that CHK1 and CHK2 could be regarded as specific antibodies against chicken BMSCs and UC-MSCs.
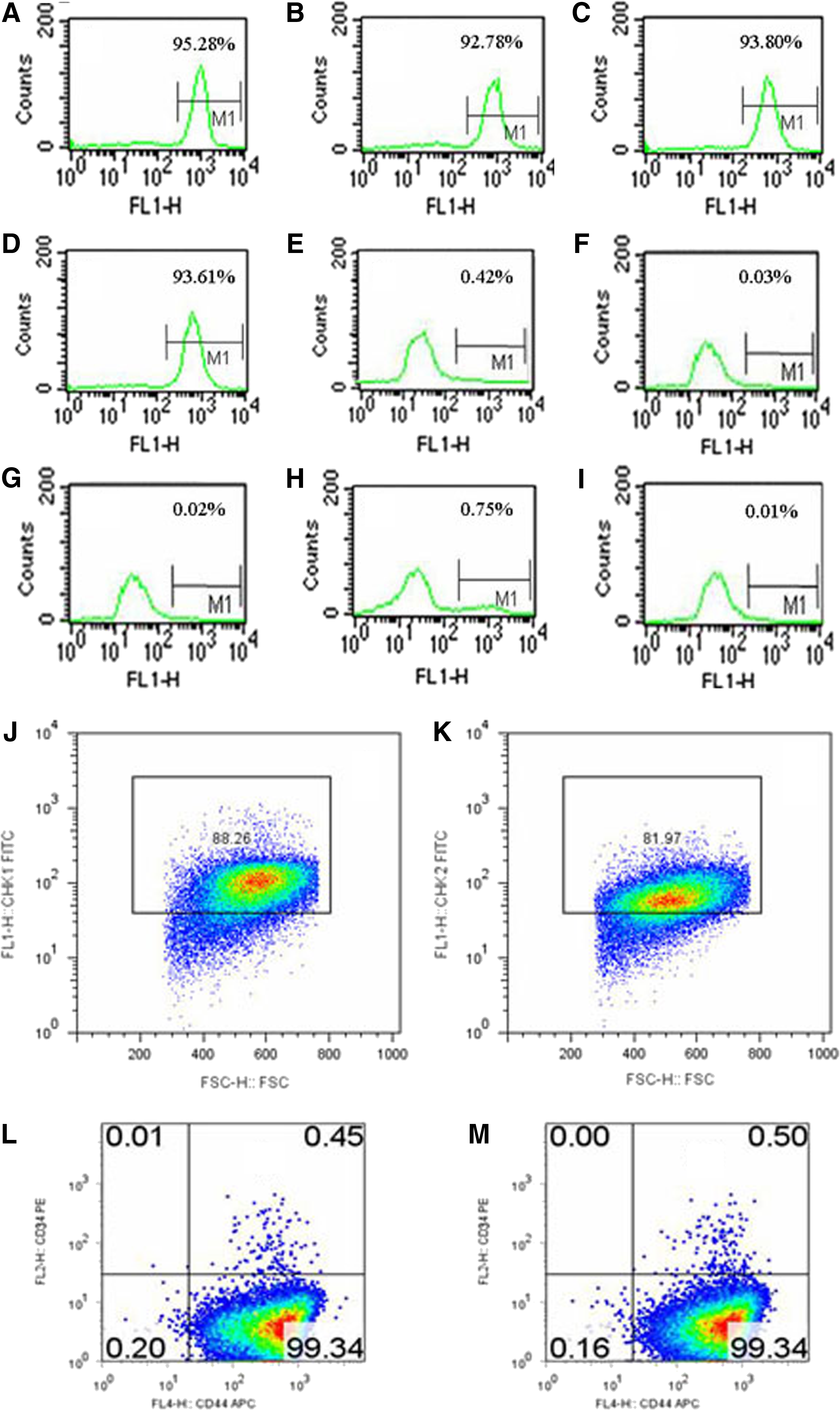
Characterization specificity of MAbs (CHK1 and CHK2) by FACS.
To evaluate the expression pattern of CHK1 and CHK2 in BMSCs, we stained BMSCs with CHK1 and CHK2. Overall, 88.26% of BMSCs displayed CHK1 expression (Fig. 6J), and 81.97% of BMSCs displayed CHK2 expression (Fig. 6K) by FACS, indicating predominant expression of CHK1 and CHK2 in BMSCs. These results indicated that CHK1 and CHK2 were cell surface proteins. A predominant expression of CD44+/CD34− was found in the CHK1+ (Fig. 6L) and CHK2+ (Fig. 6M) fraction isolated from BMSCs, respectively. These results showed that the sorted cells with two MAbs (CHK1 and CHK2) were BMSCs. The CHK1 and CHK2 can be used to develop the detection assay of the chicken BMSCs.
Immunohistochemistry
The CHK1 showed negative reactions with intestinal tissues (Fig. 7A), lung tissues (Fig. 7B), kidney tissues (Fig. 7C), heart tissues (Fig. 7D), brain tissues (Fig. 7E), and liver tissues (Fig. 7F), but it showed positive reactions with bone containing marrow (brown, Fig. 7G), bone marrow (brown, Fig. 7H), and chicken BMSCs, including the CFU-F (brown, Fig. 7I). The CHK2 showed negative reactions with intestinal tissues (Fig. 7J), lung tissues (Fig. 7K), kidney tissues (Fig. 7L), heart tissues (Fig. 7M), brain tissues (Fig. 7N), and liver tissues (Fig. 7O), but it showed positive reactions with bone containing marrow (brown, Fig. 7P), bone marrow (brown, Fig. 7Q), and chicken BMSCs, including the CFU-F (brown, Fig. 7R). These data indicated that CHK1 and CHK2 can be regarded as specific antibodies against BMSCs.
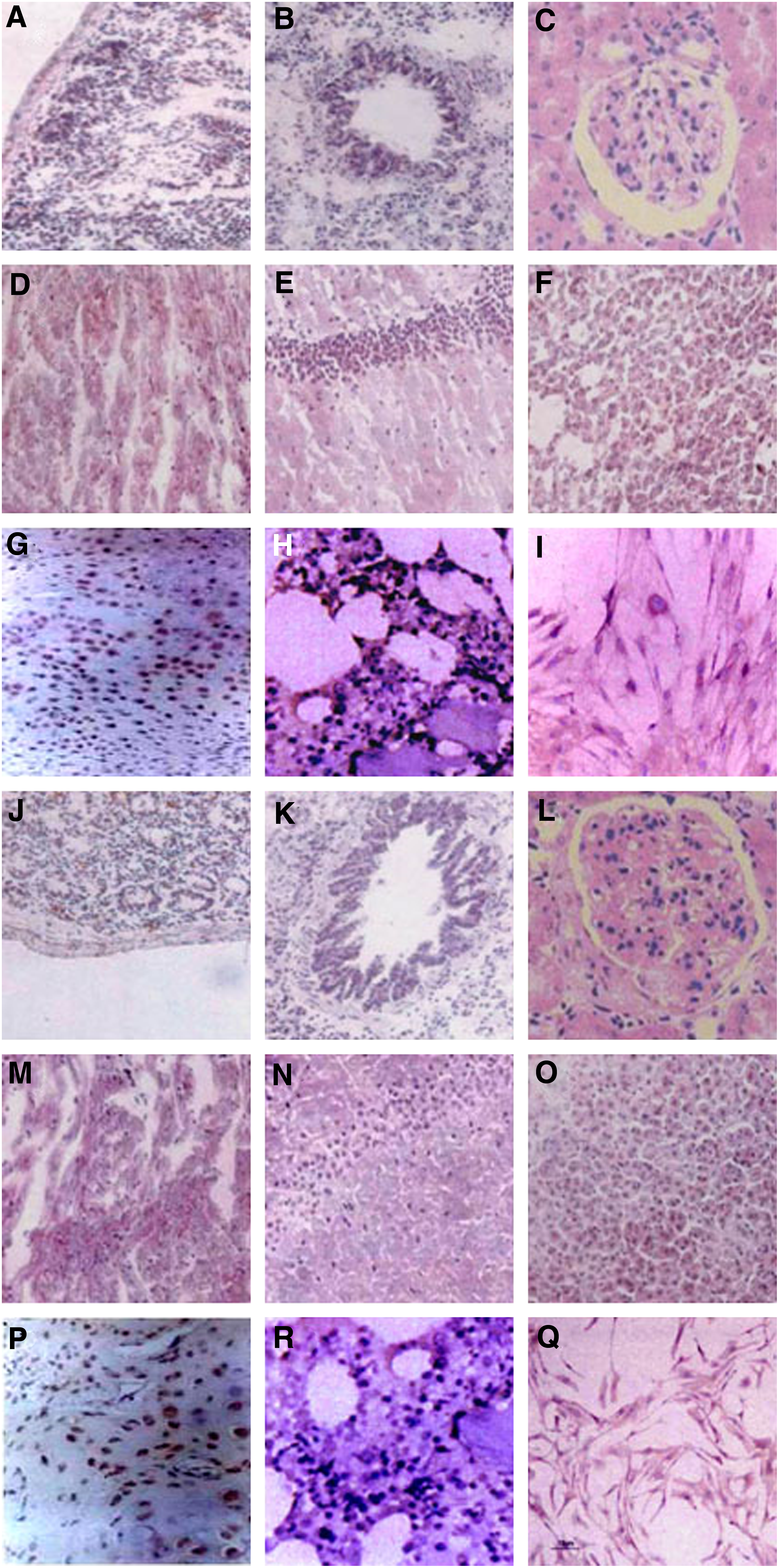
IHC analysis of MAbs (CHK1 and CHK2).
Subtype identification of MAbs
Subtype identification indicated that heavy chains of CHK1 and CHK2 were typed as IgG1 and IgG2b, respectively, and all light strands were kappa subtype.
Immunoblot reactivity of monoclonal antibody
The proteins of chicken BMSCs, chicken UC-MSCs, human BMSCs, and mouse BMSCs were separated by SDS-PAGE, respectively (Fig. 8A) and analyzed by Western blotting. CHK1 and CHK2 showed a single immunopositive band with a molecular weight of ∼90 kDa (Fig. 8B-a) and 130 kDa (Fig. 8B-b) in chicken BMSCs and chicken UC-MSCs, respectively. There was no immunopositive band in heterologous MSCs of human and mouse. These results indicated that the present CHK1 and CHK2 specifically recognized chicken BMSCs and UC-MSCs.

SDS-PAGE and Western blotting analysis of MAbs (CHK1 and CHK2).
Discussion
In the present study, we isolated and expanded adherent cells with some MSC features from chicken bone marrow. Our study detected the expression of osteogenic markers, most frequently mentioned and namely ALP activity and the initial site of mineralization,(18) and adipogenic markers, namely Oil Red O dye and PPARγ.(19,20) Our results confirmed that BMSCs could be induced to differentiate into adipogenic or osteogenic lineages by controlling microenvironments. We obtained two clones of hybridomas secreting specific antibodies directly against chicken BMSCs, namely CHK1 and CHK2.
In our experiments, the immunofluorescence assay was used to analyze the antibody production of hybridoma cells within the confocal microscopy. This procedure proved to be simple and sensitive, with which noncontaining antibody wells, weakly antibody-containing wells, and strongly antibody-containing wells could be distinguished easily because of the color development in each well. An indirect solid-phase immunoassay has been applied for anti-BMSCs hybridoma screening. Using this method, culture supernatants of growing hybridomas can be screened for antibody activity after 7 days, after the fusion experiment.
In this study, two MAbs were reactive with chicken BMSCs and UCB-MSC with a high specificity and sensitivity. With the increase of subculture, the percentage of positive cells was higher and higher, suggesting that the epitope was a fraction of large abundant bone marrow cells that survive and presumably proliferate in the adherent fraction of bone marrow. Some researchers have suggested that BMSCs are derived from bone marrow, periosteum, trabecular bone, adipose tissue, synovium, skeletal muscle, deciduous teeth, or perivascular cells.(21–25) However, our results indicated that two MAbs were nonreactive with peripheral blood, different organs, and tissues, which were not consistent with previous findings, and that reason was unclear. So, we suggested the following possible factors: (1) MSCs are a group of heterogeneous cells; whereas specific MAbs only are against one type of homogeneous cells whose populations from different tissue sources of MSCs may be different in content; (2) detection technology affects the accuracy of our results, such as reactive time, temperature, concentration of antibody, and specimen handling. For example, specimen mishandling would cause the change of morphological structure of specimens, diffusion, and loss of antigen from in situ, resulting in negative results.
On Western blotting analysis, both the MAbs showed immunopositive single bands with a molecular weight of ∼90 and 130 kDa in chicken BMSCs and chicken UC-MSCs, respectively. These results showed that they were directed against the different surface determinants present on the outer membrane. Further studies are in progress to determine the exact nature of the antigen(s) involved in the serological reaction.
Conclusion
In the present study, we isolated and expanded adherent cells with BMSCs features from chicken bone marrow, and we produced and characterized the MAbs (CHK1 and CHK2) directly against BMSCs. Both the MAbs were reactive with chicken BMSCs with a high specificity and sensitivity, and they could be used to specifically identify chicken BMSCs and UC-MSCs by ICC assays and Western blotting analysis instead of the routine means of identifying BMSCs using many markers. Subcellular location of MAbs still needs to be further lucubrated. In summary, evidence was presented that two MAbs were reactive with BMSCs, including the CFU-F. The availability of antibodies that are specific for BMSCs opened up many possibilities for further work, not only for in vitro and in vivo studies of BMSCs differentiation but also for the examination of BMSCs status in groups of patients of transplantation. In the future, populations of cells enriched in BMSCs that are prepared using antibodies may also be found in clinical application alone or in combination with biomaterials for the repair of marrow defects.
Footnotes
Acknowledgments
This study was supported by the National Natural Science Foundation (30873190, 31300660, 81402287), the science and technology innovation action plan of Shanghai (14431904300), Shanghai Pujiang Program (13PJD012), and the foundation for young teacher from Education Ministry of China (20120074120027 and 20130073-120010) and it was partially supported by the Open Funding Project of the State Key Laboratory of Bioreactor Engineering.
Author Disclosure Statement
No competing financial interests exist.