
Abstract
The bone marrow stromal cell antigen 2 (BST-2) protein was identified as a novel virus restriction factor that potently restricts the replication and egress of enveloped viruses. In this study, we generated monoclonal antibodies (MAbs) against porcine BST-2 encoding 34–112 aa of porcine BST-2, which was cloned and inserted into the prokaryotic expression vector pCold-I to construct a recombinant plasmid pCold-pBST-2. The recombinant porcine BST-2 protein (rpBST-2 protein) was induced by isopropyl-β-D-thiogalactoside in Escherichia coli BL21 (DE3). Then, BALB/c mice were immunized with the purified rpBST-2 protein to prepare MAbs of BST-2. After subcloning, one strain of hybridoma cells named 1B2 secreting porcine BST-2 protein monoclonal antibody (MAb) was obtained. Indirect immunofluorescence assay and western blot analysis showed that the MAb was specifically reacted with the overexpressed porcine BST-2 protein in Vero cells. The specific MAb of porcine BST-2 provides a valuable tool for further studies of BST-2 to restrict virus infection.
Introduction
T
BST-2 inhibits the release of enveloped viruses at the plasma membrane (PM) and is a broad antiviral protein against enveloped viruses such as HIV, simian immunodeficiency virus (SIV), Lassa and Marburg virus, and Ebola virus-like particles.(6–8) Some studies showed BST-2 inhibits hepatitis C virus (HCV), hepatitis B virus, dengue virus, and human parainfluenza virus type 2.(9–12) Recently, BST-2 was identified as a pattern recognition receptor to activate NF-κB in response to viral tethering and acted as an innate sensor of viral assembly.(13,14)
To facilitate the study of the antiviral activity of BST-2 to enveloped virus infection, we used the recombinant porcine BST-2 protein (rpBST-2 protein) to prepare monoclonal antibodies (MAbs) against porcine BST-2, which are the preferred antibodies for investigating the functions of BST-2.
Materials and Methods
Cells, antibodies, and animals
SP2/0 myeloma cells and African green monkey kidney cells (Vero E6) were obtained from Shanghai Cell Biology Institutes, Chinese Academy of Sciences (Shanghai, China). Dulbecco modified Eagle's medium, Opti-MEM cell culture medium, and fetal bovine serum were purchased from Gibco (Grand Island, NY) at 37°C in a 5% CO2 atmosphere. Anti-FLAG antibody, anti-β-actin antibody, Alexa Fluor 488 donkey anti-mouse IgG (H + L) antibody, horseradish peroxidase (HRP)-conjugated goat anti-mouse, and p3×FLAG-CMV-7.1 plasmids were purchased from Sigma (Shanghai, China). The 6-week-old female BALB/c mice were purchased from the Shanghai Slack Laboratory Animal (Shanghai, China).
Expression and purification of rpBST-2 protein
According to the BST-2 nucleotide sequence of Sus scrofa (GenBank no. AK346678), a pair of primers (forward primer 5′-TACCCTCGAG
Immunization
Purified rpBST-2 protein (50 mg/mouse) was emulsified with an equal amount of Freund's complete adjuvant and subcutaneously injected into BALB/c mice. This was followed by three additional injections at 2-week intervals for each mouse with the same dose of the antigen plus Freund's incomplete adjuvant. Three days after the last immunization, spleen cells of immunized mice were isolated and fused with mouse SP2/0 myeloma cells.
Preparation of anti-rpBST-2 protein-specific monoclonal antibody
The anti-rpBST-2 protein serum was obtained from the immunized mice and the antibody titers were determined using indirect ELISA. Spleen cells from the immunized mice with the highest anti-pBST-2 serum antibody titer were fused with SP2/0 myeloma cells under the action of 50% PEG as fusion agents. The hybridoma cells were cultured in 96-well plates in HAT screening culture medium at 37°C in a humidified 5% CO2 incubator. Indirect ELISA was used to filter the positive hybridomas when the cells had covered one third of the bottoms of 96-well plates. After cloning four times by limiting dilution, the positive hybridoma cells were injected into pristane-treated BALB/c mice to gain abundant ascetic fluid.
Indirect enzyme-linked immunosorbent assay
Indirect ELISA was used to determine the screen of positive hybridoma cells and antibody titer. The ELISA plates were plated with 100 mL purified rpBST-2 protein (200 ng/well) in carbonate bicarbonate buffer (15 mM Na2CO3, 35 mM NaHCO3 [pH 9.6]) and coated at 4°C overnight. The plates were then blocked with 5% skimmed milk in phosphate buffer with 0.05% Tween-20 (PBST) at 37°C for 1 hour. After washing thrice, the plates were incubated with 100 μL diluted cell culture supernatant or antibodies at 37°C for 1 hour. The plates were incubated with HRP-conjugated goat anti-mouse IgG with 1:10,000 dilution in PBST at 37°C for 1 hour after washing thrice in PBST. Then, the plates were incubated with 50 μL/well of TMB liquid (Amresco, Solon, OH) for 15 minutes at room temperature with protection from light. The results were read with an OD450 value after being stopped by 2 M H2SO4. The wells coated with purified His-tag protein served as negative control for screening the positive hybridoma cells.
Eukaryotic expression vector construction and transfection
According to the full-length BST-2 nucleotide sequence of Sus scrofa (GenBank no. AK346678), a pair of primers (forward primer 5′-CGACTCTAGA
For transfection, Vero cells were seeded in six-well plates and transfected at 80% confluency with pFLAG-pBST-2 plasmids DNA in Gibco Opti-MEM cell culture medium by FuGENE HD Transfection Reagent (Promega, Madison, WI), according to the manufacturer's instructions. Empty vector transfection served as control. After 24 hours transfection, the cells were harvested and detected by Western blot analysis.
Western blot analysis
Specificity of the MAbs was confirmed by Western blot analysis. The cells' protein was collected and separated by 10% SDS-PAGE, then transferred to the nitrocellulose (NC) membrane. The membrane was blocked with 5% skimmed milk in TBST (TBS with 0.1% Polysorbate-20) for 1 hour at room temperature. The anti-FLAG antibody, anti-β-actin antibody, or anti-BST-2 MAbs were added on the NC membrane for 1 hour at room temperature. After washing thrice with TBST, the membrane was incubated with the HRP-conjugated goat anti-mouse IgG (1:6000 dilution in TBST) for 1 hour at room temperature. After washing, the SuperSignal West Pico chemiluminescent substrate (Thermo Fisher Scientific, Waltham, MA) was used for color development.
Indirect immunofluorescence assays
Vero cells were plated in a six-well plate and fixed by paraformaldehyde for 30 minutes at room temperature at 80% cell confluency. After washing twice in phosphate-buffered saline (PBS), cells were permeabilized by incubation with 0.25% Triton-100 in PBS for 5 minutes. The cells were blocked with 10% bovine serum albumin in PBS for 1 hour at 37°C after washing twice in PBS. The cells were then incubated with the anti-BST-2 monoclonal antibody (MAb) in PBS at 37°C for 1 hour. This was followed by washing four times in PBS; the cells were then incubated with Alexa Fluor 488 donkey anti-mouse IgG (H + L) antibody at 37°C for 1 hour. The cells were stained with DAPI for 5 minutes at 37°C, followed by washing twice. The cells were washed four times in PBS and then observed under fluorescence microscope.
Results
Expression of rpBST-2 protein
The total RNA was extracted from small intestine tissue of piglet and was used for cDNA synthesis. The 336 bp fragment and the full length of porcine BST-2 gene were amplified (Fig. 1) and inserted into the expression vector. Sequencing results show that the BST-2 gene inserted into the correct position. The expression vector pCold-I containing 336 bp fragment of porcine BST-2 gene was induced by IPTG, and the high-level expression of soluble from rpBST-2 was achieved. The recombinant protein (∼13 kDa) was easily purified by His-binding resin and confirmed by SDS-PAGE and Western blot analysis. The empty vector transformed bacteria served as control (Fig. 2).
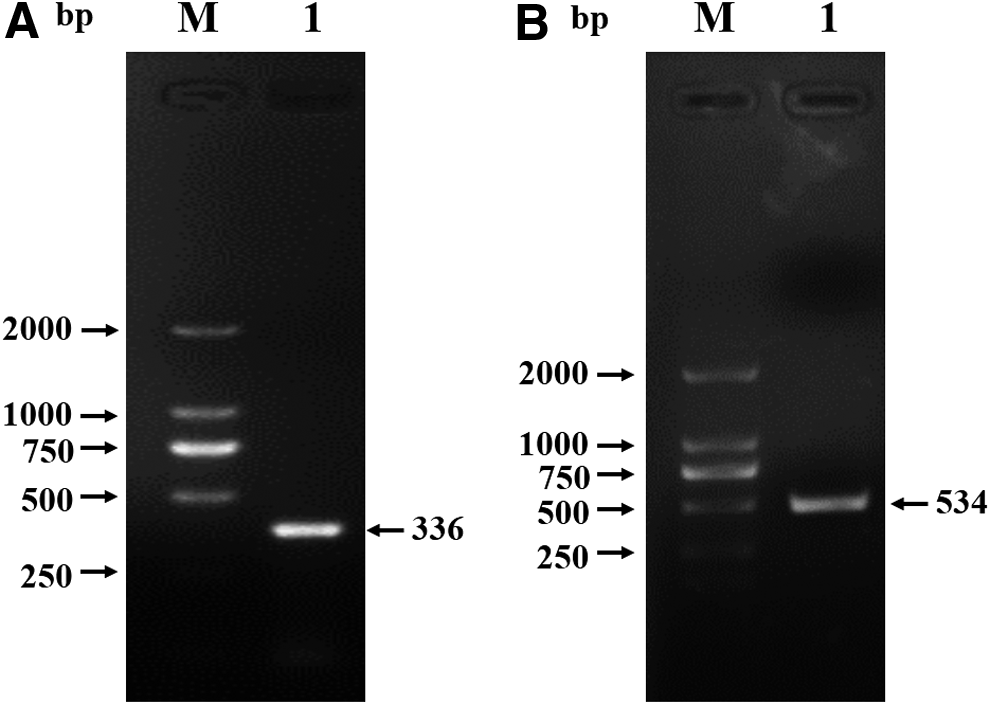
Amplification of porcine BST-2 gene. Extracellular domain
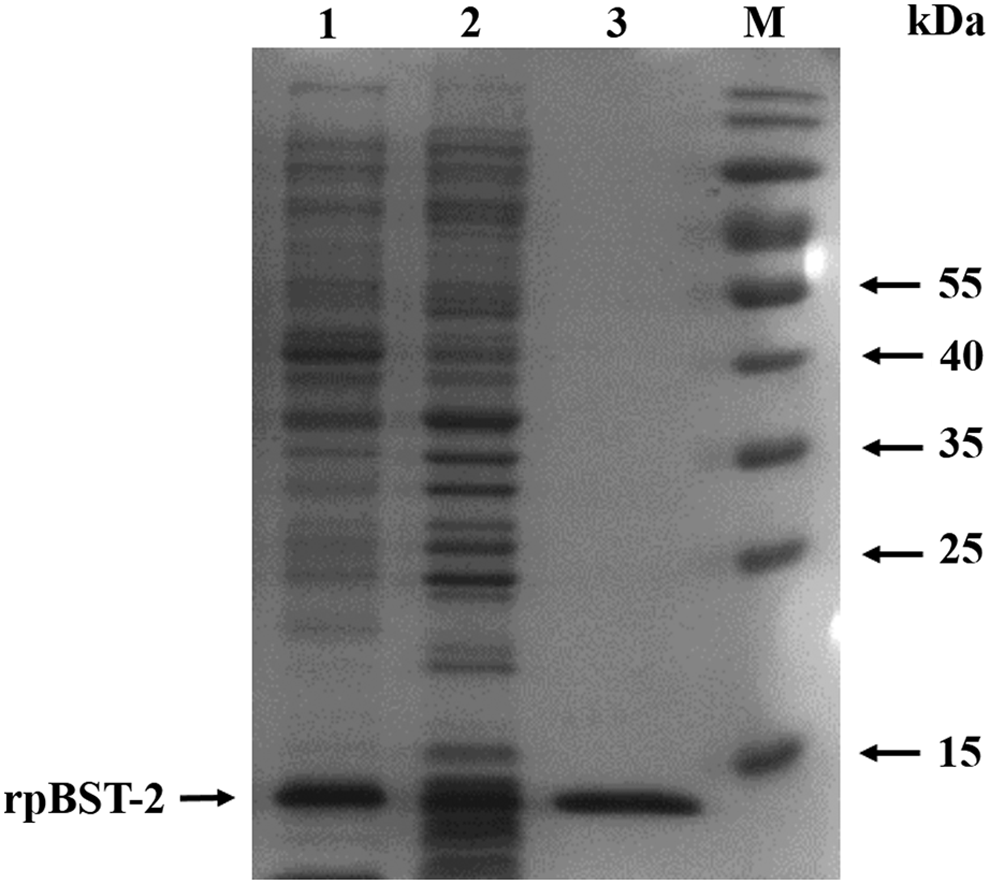
Expression and purification of recombinant porcine BST-2 protein analyzed by SDS-PAGE. Lane 1, supernatant proteins; lane 2, precipitation proteins; lane 3, purified rpBST-2 protein; M, PageRuler prestained protein ladder. SDS-PAGE, sodium dodecyl sulfate-polyacrylamide gel electrophoresis.
Generation of porcine BST-2 MAbs
The purified rpBST-2 protein was used to immunize BALB/c mice to prepare MAbs. The antisera titers were determined by indirect ELISA after four immunizations. The mouse with the highest antibody titers against rBST-2 was used for cell fusion. After subcloning four times by limiting dilution and screening, one positive MAb against porcine BST-2 protein was identified and named 1B2.
Reactivity of MAb 1B2
The MAb 1B2 cell clone was used to prepare the ascites containing MAbs. The ascites were purified using NAb™ Protein G Spin Columns (Thermo Fisher Scientific, Rockford, IL). Western blot analysis (Fig. 3) and Immunofluorescence assay (IFA) (Fig. 4) were used to identify the specificity of the MAbs against overexpressed porcine BST-2 protein in Vero cells. The results suggested that the MAb could be specifically reacted with porcine BST-2 protein.
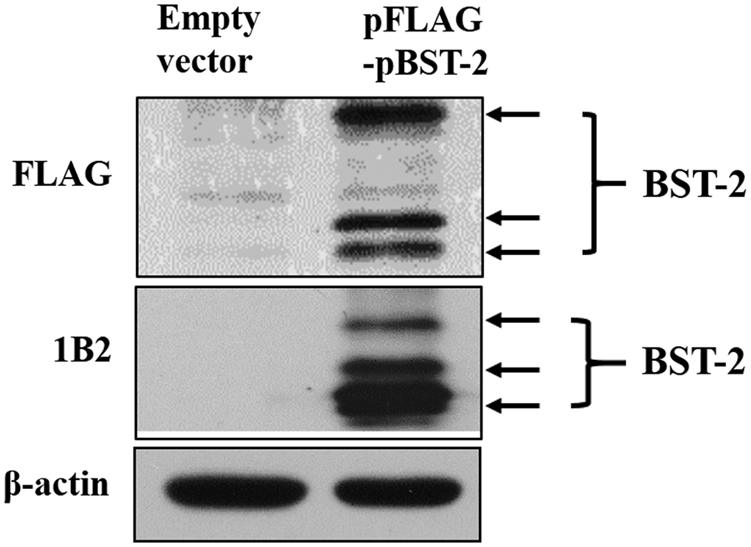
Western blot analyzed the specificity of prepared MAbs against porcine BST-2. Expression of overexpressed porcine BST-2 in Vero cells was detected by anti-FLAG antibody, BST-2 MAb 1B2, and anti-β-actin antibody. The protein of β-actin was detected as a control. All the experiments were repeated thrice and representative results were shown. MAb, monoclonal antibody; MAbs, monoclonal antibodies.

Indirect immunofluorescence assays analyzed the specificity of prepared MAbs against porcine BST-2. Vero cells were plated and transfected in six-well plates. Twenty-four hours later, cells were incubated with MAb 1B2 or normal mouse antibody (negative control), followed by Alexa Fluor 488 donkey anti-mouse IgG (H + L) antibody.
Discussion
BST-2 is a transmembrane protein that localized at the PM, as well as the membranes of multiple intracellular vesicles,(15) and is a pattern recognition receptor to activate NF-κB in response to viral tethering and acted as an innate sensor of viral assembly.(13,14) The expression of BST-2 is induced by IFN-α, -β, -γ, -τ, -λ3, and–ω in many organs and tissues(16–21) and which is upregulated in viral infection.(22–24) BST-2 is a broad antiviral protein inhibiting the release of viruses, such as HIV, SIV, Lassa and Marburg virus, Ebola virus-like particles,(6–8) HCV, Hepatitis B Virus, dengue virus, and Human parainfluenza virus type 2.(9–12)
Porcine epidemic diarrhea virus, highly pathogenic porcine reproductive and respiratory syndrome virus, porcine pseudorabies virus, classic swine fever virus, swine Japanese encephalitis virus, and other viruses pose a serious threat to the swine industry worldwide. The relationship between BST-2 and virus infection is not well determined and there are no specific MAbs against porcine BST-2 that could be used. In this study, the porcine BST-2 gene was expressed bacterially for generation of MAbs against porcine BST-2. A high level of antibody induced by the purified rBST-2 protein was detected using indirect ELISA in immunized mice. Following hybridoma technique, a MAb (1B2) against porcine BST-2 was generated and identified by Western blot and IFA. This antibody provided a valuable tool for further investigation of the antiviral activity of BST-2.
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of China (grant no. 31201915) and the Shanghai Science and Technology Program for Agriculture (grant nos. 2013-5-5 and 2013-3-6).
Author Disclosure Statement
No competing financial interests exist.