
Abstract
Immunoglobulin A (IgA) antibodies exist in monomeric, dimeric, and secretory forms. Dimerization of IgA depends on a 15-kD polypeptide termed “joining (J) chain,” which is also part of the binding site for an epithelial glycoprotein called “secretory component (SC),” whether this after apical cleavage on secretory epithelia is ligand bound in secretory IgA (SIgA) or in a free form. Uncleaved membrane SC, also called the “polymeric Ig receptor,” is thus crucial for transcytotic export of SIgA to mucosal surfaces, where it interacts with and modulates commensal bacteria and mediates protective immune responses against exogenous pathogens. To evaluate different forms of IgA, we have produced mouse monoclonal antibodies (MAbs) against human J-chain and free SC. We found that J-chain MAb 9A8 and SC MAb 9H7 identified human dimeric IgA and SIgA in enzyme-linked immunoassay and western blot analysis, as well as functioning in immunohistochemistry to identify cytoplasmic IgA of intestinal lamina propria plasmablasts/plasma cells and crypt epithelium of distal human intestine. Finally, we demonstrated that SC MAb 9H7 cross-reacted with rhesus macaque SIgA. These novel reagents should be of use in the study of the biology of various forms of IgA in humans and SIgA in macaques, as well as in monitoring the production and/or isolation of these forms of IgA.
Introduction
I
Mouse monoclonal antibodies (MAbs) for detecting dIgA and SIgA are of interest to define the roles of different forms of IgA in plasma and mucosal secretions and monitoring isolation or production of dIgA and SIgA. In this study, we report the production of two new murine MAbs, 9A8 against human J-chain and 9H7 against human SC, and their use in characterizing recombinant IgA molecules.
Materials and Methods
Genes and recombinant proteins of J-chain, SC, and IgA antibodies
DNA sequences of human J-chain (accession number: P01591)(30) and pIgR of human (accession number: NM_002644)(31) and rhesus macaque (accession number: EHH15602)(31) were obtained from the GenBank. For production of soluble recombinant proteins and use as DNA immunization in mice, genes encoding for human J-chain and human and rhesus free SC(2,32) were codon optimized by converting amino acid sequences to nucleotide sequences using the codon usage of highly expressed human housekeeping genes and de novo synthesized (GenScript). All constructs contained the C-terminal His6 tag for purification by nickel columns. The synthesized gene constructs were subcloned into mammalian expression vector pcDNA3.1+ (Life Technologies) for production of soluble recombinant proteins and also for use as DNA immunogens in mice for generation of MAbs. Recombinant soluble J-chain and SC proteins were produced in 293F cells (Life Technologies) by transient transfection and purified by nickel columns with methods described previously.(33)
Mammalian expression vector pOptiVEC-TOPO was obtained from Life Technologies (catalog no. 12744-017) for producing stable CHO cell lines. The original pOptiVEC-TOPO plasmid contains selection marker and allows gene copy amplification under methotrexate hydrate drug selection once the gene insert is integrated into the host genome for effectively selecting transfectants with high copy numbers of the integrated target gene(34,35) and was used to express human J-chain gene alone (Supplementary Fig. S1a). The vector was further modified to accommodate dual expression of both J-chain and SC genes under control of two human CMV promoters in a single vector (Supplementary Fig. S1b).
Production of mIgA, dIgA, and SIgA
Gene sequences of the constant region of human IgA1 (accession number: BC092449)(36) and IgA2 (accession number: BC073765)(36) were obtained from the GenBank, de novo synthesized (GenScript), and subcloned into mammalian expression vector pcDNA3.1+ (Life Technologies). Sequences of the variable region of heavy chain (VH) of the HIV-1 broadly neutralizing IgG1 antibody (CH31) targeting the HIV-1 gp120 CD4-binding site (CD4bs),(37,38) a nonneutralizing antibody (7B2) targeting HIV-1 Env gp41 immunodominant region,(39) and an influenza A H1 subtypic broadly neutralizing antibody (CH65) targeting influenza hemagglutinin receptor bs(40,41) were selected as model systems for production of IgA antibodies and cloned into vector expressing either the constant region gene of human IgA1 or IgA2. Dimeric IgA antibodies were produced in the J-chain stably transfected CHO cells by cotransfection with CH31 or 7B2 IgA1 or IgA2 heavy chain plasmid and their corresponding light chain plasmid and purified by peptide M/Agarose beads (catalog no. gel-pdm-5; InvivoGen) according to the protocol recommended by the manufacturer. Production of CH31 dIgA2 and 7B2 dIgA2 in hundreds of mg scale was carried out by Catalent Pharma Solutions in CHO cells with the same human J-chain, CH31 IgA2 and 7B2 IgA2, heavy and light chain gene constructs, and KappaSelect beads (GE Healthcare Life Science) for IgA purification. It was reported that SIgA could be produced via copurification process of dimeric IgA containing J-chain with recombinant human free SC.(42) Two approaches were used to produce CH31 SIgA antibodies. First, advantage was taken of the ability of the dIgA antibody to bind free SC by mixing and incubation.(42) Thus, CH31 SIgA antibodies were produced by complexing purified CH31 dIgA with recombinant free SC at 4° overnight at a 1:1 molar ratio, followed by purification with size exclusion column chromatography. Second, CH31 SIgA antibodies were produced in the J-chain and SC stably transfected CHO cell lines by transfection with CH31 IgA2 heavy and light chain plasmids and purified by Peptide M/Agarose beads (catalog no. gel-pdm-5; InvivoGen), as recommended by the manufacturer.
J-chain antibody production
BALB/c mice were immunized intramuscularly four times with 50 μg of human J-chain expressing plasmid DNA in saline, followed by one boost with the purified recombinant J-chain and Sigma Adjuvant System (catalog no. S6322-1VL; Sigma-Aldrich, Inc.) 3 days before harvesting for fusion. The spleen was harvested, and 1.56 × 107 splenocytes were fused to NS0 Bcl2 cells(43,44) with Polyethylene Glycol 1500 (catalog no. 10 783 641 001; Roche) by methods described previously.(43,45) A total of 1632 wells were plated in SuperHAT media at a density of ∼10,000 cells/well and cultured for 14 days prior to harvesting culture supernatants for antibody screening.(45) The collected culture supernatants were screened against the purified recombinant human J-chain protein by enzyme-linked immunoassay (ELISA). Two human J-chain-reactive hybridoma cell lines, 4G10 and 9A8, were identified and cloned by limiting dilution, adapted to culture condition with serum-free media (catalog no. 12045-076; Gibco), and expanded in CELLine bioreactors (catalog no. WCL1000-1; Wheaton) for production of MAbs. J-chain MAbs 4G10 and 9A8 were purified from the culture supernatants by HiTrap Protein G HP columns (catalog no. 17-0404-01; GE Life Sciences).
SC antibody production
BALB/c mice were immunized intramuscularly four times with 50 μg of human SC-expressing plasmid DNA in saline, followed by one boost of recombinant SC protein with Sigma Adjuvant System. A subsequent boost with recombinant protein was performed 3 days before fusion. Approximately 1.44 × 107 splenocytes were collected from the harvested spleen and fused to NS0 Bcl2 cells by Polyethylene Glycol 1500. A total of 1440 wells were plated in SuperHAT media at a density of ∼10,000 cells/well and expanded for 14 days. The culture supernatants were collected from the individual culture wells and screened against recombinant free SC by ELISA. Hybridoma clone 9H7 was identified and cloned by limiting dilution. Clonal 9H7 cell line was adapted to culture condition with serum-free media and expanded in CELLine bioreactors. SC MAb 9H7 was purified from the culture supernatants by HiTrap Protein G HP columns.
Immunohistology
Resection preparation of the distal human ileum without histological signs of pathology was used. It was fixed in formalin, and serial sections were cut at 6 μm, dewaxed, and pretreated with citrate buffer to unmask antigenic determinants before immunostaining. Various concentrations of the primary antibodies were tested to select optimal conditions, and standardized secondary goat anti-mouse or anti-rabbit IgG reagent—labeled with either peroxidase or Alexa Fluor (R) (Molecular Probes; Thermo Fisher Scientific)—was then applied. As reference primary antibody, our rabbit R-282(46) raised against human J-chain was used, and a rabbit antibody against human SC was applied as the purified immunoglobulin fraction (DAKO).
Animal experiments and human samples
Work related to mouse immunization and sampling and collection of rhesus blood samples was approved by the Duke Institutional Animal Care and Use Committee. Human breast milk (BM) and serum samples were collected in compliance with Institutional Review Board protocols approved by the Duke University Health System Institutional Review Board. Human intestinal resection preparation of the distal ileum was from collection of biological material that was intended for clinical examination, diagnostics, and treatment and does not require explicit informed consent according to The Norwegian Treatment Biobank Act 2003.
Enzyme-linked immunoassay
Reactivity of CH31 and 7B2 antibodies in forms of IgA1, IgA2, mIgA, dIgA, and SIgA as well as IgG with HIV-1 Env CON-S gp120 and HIV-1 Env gp41 immunodominant epitope peptide SP400 (RVLAVERYLRDQQLLIWGCSGKLICTTAVPWNASWSNKSLNKI) was tested in standard sandwich ELISA. HIV-1 Env CON-S gp120 or HIV-1 Env gp41 SP400 peptide was coated at saturated amounts (2 μg/mL and 30 ng/well) in 384-well ELISA plates overnight at 4°C, followed by incubation with serial dilutions (from 100 to 0.001 μg/mL) of CH31 or 7B2 in forms of mIgA1, mIgA2, and dIgA, with IgG antibody as control. To measure the binding of free SC to dimeric IgA antibodies, CH31and 7B2 dIgA were coated at saturated amounts (2 μg/mL and 30 ng/well), with mIgA antibody as control, in 384-well ELISA plates overnight with either human or rhesus free SC at concentrations ranging from 50 μg to 0.023 μg/mL 4°C, followed by incubation with SC MAb 9H7. To determine binding of J-chain MAb 9A8 to dimeric IgA, CH31 dIgA2 was coated at saturated amounts (2 μg/mL and 30 ng/well) in 384-well ELISA plates overnight at 4°C, followed by incubation with J-chain MAb 9A8 at concentrations ranging from 100 μg to 0.01 μg/mL. After incubation, these plates were washed twice with PBS and then incubated with secondary antibody of horseradish peroxidase (HRP)-conjugated goat anti-human IgA (Jackson Immunoresearch) (1/5000 dilution in 5% goat serum/PBS), goat anti-human IgA IgG, or goat anti-mouse IgG accordingly. These plates were then washed four times with PBS and developed with 30 μL/well TMB substrate (SureBlue Reserve). The HRP reaction was stopped with 30 μL/well 1 M HCl, and optical density (OD) of the reaction was measured at 450 nm.
Sodium dodecyl sulfate–polyacrylamide gel electrophoresis, blue native (BN)-PAGE, and western blot analysis of the Env proteins
Purified J-chain, free SC, and CH31 IgA antibodies purified from culture supernatants of the transfected 293F cells or plasma or milk were fractionated at 2 μg/lane on 4%–20% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) gels (Invitrogen) under reducing (4% beta-mercaptoethanol, β-ME; Fisher Scientific) or nonreducing conditions, and IgA antibodies (HG129 and HG130) purified from EBV-transformed human B cell lines were fractionated at 4 μg/lane on 4%–20% BN-PAGE.(47) After electrophoresis, gels were either stained with Coomassie blue or transferred onto nitrocellulose filters and probed with anti-J-chain mouse MAbs (4G10 and 9A8), or rabbit antibody to J-chain (R-282) or anti-human IgA antibody. Secondary antibodies of goat anti-mouse or anti-rabbit IgG–alkaline phosphatase conjugates (Sigma Aldrich, Inc.; 1/3000 dilution) were used, and the western blots were developed with Western Blue Substrate (Promega).
Results
Production of monomeric, dimeric, and secretory forms of IgA
Two human HIV-1 MAbs, CH31 (an HIV-1 broadly neutralizing antibody targeting HIV-1 gp120 CD4-bs)(37,38) and 7B2 (a nonneutralizing and ADCC-mediating antibody targeting HIV-1 Env gp41 immunodominant region),(39) were produced as purified mIgA1, mIgA2, and IgG1 as control and assayed for their ability to bind HIV-1 Env antigens (Fig. 1). When tested for binding to HIV-1 CON-S gp140 Env protein(48) in ELISA, both purified CH31 mIgA1 and mIgA2 bound CON-S gp140 Env protein with similar EC50 values compared to CH31 IgG1 antibody (Fig. 1a). 7B2 mIgA2 and IgG1 antibodies bound to HIV Env gp41 with similar apparent binding affinities (Fig. 1b), while 7B2 mIgA1 had weaker binding to HIV Env gp41 with EC50 of 0.05 μg/mL compared with the binding by 7B2 IgG with EC50 of 0.006 μg/mL (Fig. 1b). Each of the IgA forms of CH31 showed potent neutralization of two tier 2 (neutralization resistant) HIV-1 pseudoviruses (A.Q23 and A.Q168) (Fig. 1c). However, compared to CH31 IgG for neutralization activity, the 50% inhibitory concentration (IC50) values were weaker for mIgA2 than for mIgA1 (Fig. 1c). Because human IgA2 is generally more resistant to protease cleavage than IgA1 antibodies,(1,23,49) human IgA2 was selected as a model to generate dIgA and SIgA for further studies.
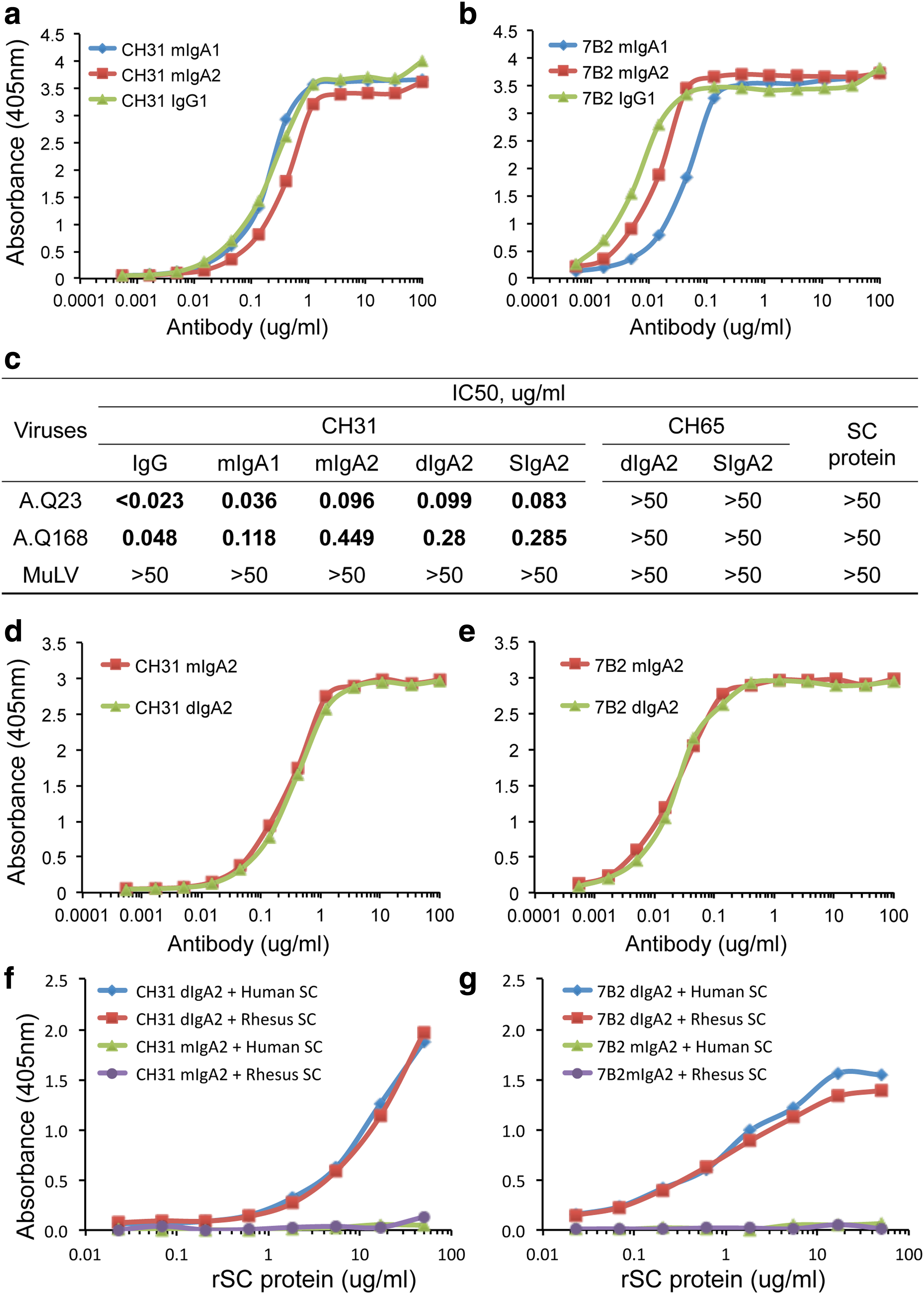
Comparison of binding and neutralizing activity of mIgA1, mIgA2, IgG, dIgA2, and SIgA antibodies. Serial dilutions from 100 μg/mL to 0.001 μg/mL of purified CH31
We next asked if there were differences in the binding ability of mIgA2, dIgA2, and SIgA2 antibodies. We found that there were no differences between mIgA2 and dIgA2 antibodies in binding of either CH31 (Fig. 1d) or 7B2 (Fig. 1e) to HIV-1 Env or neutralization activity by CH31 (Fig. 1c). To produce SIgA antibodies, we first determined if dIgA2 antibodies could bind directly to free SC protein in vitro. Both CH31 dIgA2 and 7B2 dIgA2 were tested for binding to both human and rhesus free SC. We found that dIgA2 antibodies of both CH31 (Fig. 1f) and 7B2 (Fig. 1g) bound both to human free SC and to recombinant rhesus free SC, while mIgA2 antibodies of CH31 and 7B2 containing no J-chain did not bind to either human or rhesus SC (Fig. 1f, g). Interestingly, dIgA2 antibodies with different specificities (CD4 bs vs. gp41) had quite different binding slopes to recombinant SC proteins in keeping with the fact that antibody variable region plays a role in the binding of SC to dIgA antibodies.(1) We also found that CH31 SIgA2 produced in vitro by incubation of SC with CH31 dIgA2 manifested the same neutralization potency in the TZMbl HIV-1 neutralization assay as mIgA2 or dIgA2 antibodies (Fig. 1c).
J-chain MAb characterization
Initial hybridoma supernatant assays identified both 4G10 and 9A8 hybridoma B-cell clones reactive with the recombinant J-chain protein. We found that MAb 4G10 was IgG2b with kappa light chain, while MAb 9A8 was IgG1 with kappa light chain. Purified MAbs 4G10 and 9A8 were tested for binding to recombinant J-chain protein and CH31 dIgA containing J-chain in western blots and ELISA with our rabbit anti-human J-chain R-282(46) as a positive control (Fig. 2). We found that the J-chain MAb 4G10 bound to recombinant J-chain protein in western blots under reducing and nonreducing conditions (Fig. 2a, b) as well as ELISA (Fig. 2e, f) but did not bind to J-chain associated with CH31 dIgA2 (Fig. 2a, b, and f). In contrast, we found that J-chain MAb 9A8-like R-282 could detect recombinant J-chain as monomers, dimers, and oligomers in nonreducing SDS-PAGE (Fig. 2a) and predominantly monomer with a minor fraction of J-chain dimers in reducing SDS-PAGE (Fig. 2b). MAb 9A8 also detected CH31 dIgA2 containing J-chain in nonreducing SDS-PAGE (Fig. 2a). Compared to our rabbit anti-J-chain R-282, MAb 9A8 showed an identical detection pattern of CH31 dIgA in SDS-PAGE under nonreducing (Fig. 2a) and reducing (Fig. 2b) conditions and, importantly, could also detect human natural dIgA purified from IgA-secreting human B-cell lines (Fig. 2c, d). J-chain MAb 9A8 was reactive with both the recombinant J-chain and the dIgA2 containing J-chain (Fig. 2e, f) in ELISA. These results demonstrated that MAb 9A8 could potentially be used to quantitate polymeric IgA in human specimens by ELISA. Neither MAbs 9A8 nor 4G10 bound to rhesus recombinant J-chain (not shown).
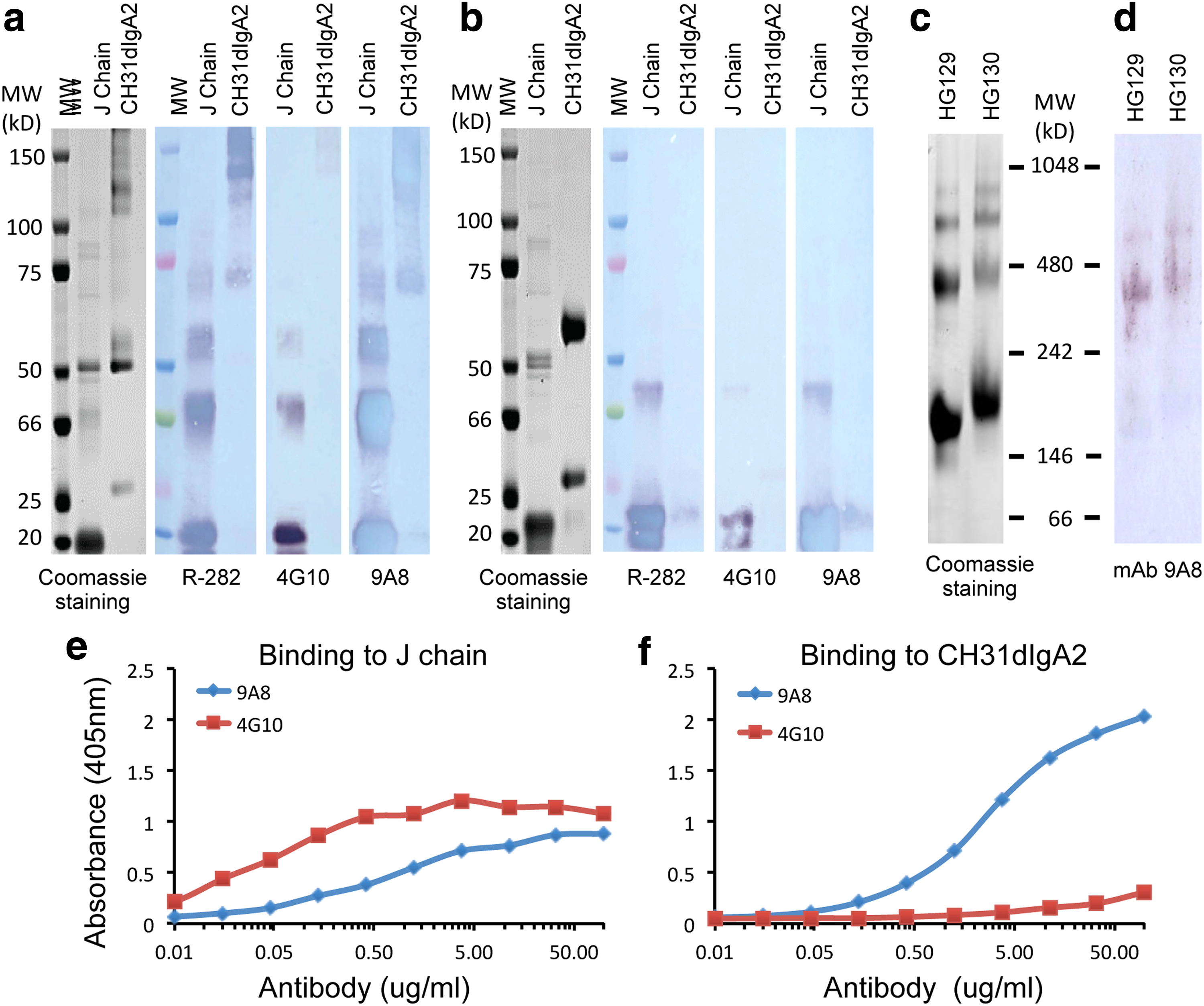
Reactivity of J-chain antibodies to recombinant J-chain, CH31 dIgA2, and human IgA MAbs derived from human B cell lines. Recombinant J-chain and purified CH31 dIgA2 were fractionated in 4%–20% gradient SDS-PAGE under nonreducing
SC MAb characterization
Culture supernatants of hybridoma cells derived from human recombinant (r) free SC-immunized mice were screened for SC-specific antibodies by ELISA against free rSC protein. One hybridoma cell line 9H7 with strong binding to the rSC protein was identified. Purified MAb 9H7 was found to be IgG2a isotype with a kappa light chain and to bind free rSC in western blots under reducing and nonreducing conditions (Supplementary Fig. S2). Using CH31 dIgA (Fig. 3a) that did not contain SC as a negative control, and free rSC as a positive control, we tested MAb 9H7 for binding to CH31 SIgA2 (Fig. 3b), which was produced by incubation of purified CH31 dIgA2 with free rSC, followed by size exclusion chromatography to remove free rSC. We found that both purified CH31 dIgA (Fig. 3a) and in vitro-formed CH31 SIgA (Fig. 3b) appeared as a single major peak in size exclusion chromatography. However, due to the denaturing conditions of SDS-PAGE, both CH31 dIgA and CH31 SIgA migrated both as whole dimers and as fragments containing monomers of heavy and light chains (Fig. 3c), according to what has been reported for recombinant IgG in SDS-PAGE.(50) MAb 9H7 detected no CH31 dIgA that contained only J-chain and no SC but did detect CH31 SIgA as high-molecular weight protein bands (150 kD and larger) and a 75-kD free rSC band (Fig. 3d).

Detection of SIgA by western blots with anti-SC MAb 9H7. CH31 SIgA antibody was produced by either in vitro formation by incubation of CH31 dIgA with free SC
We next tested MAb 9H7 for binding to CH31 SIgA2 antibody produced from J-chain and free SC transfected CHO cell line (Fig. 3e). CH31 dIgA was produced with J-chain-alone stably transfected CHO cell line and used as control (Fig. 3e). J-chain MAb 9A8 detected both CH31 dIgA and CH31 SIgA that migrated as whole antibody dimers as well as fragments containing heavy and light chain monomers with associated J-chain in nonreducing SDS-PAGE (Fig. 3f), while J-chain MAb 9A8 detected free J-chain (∼20 kD) and also J-chain in the presence of SC (∼95 kD) in the reducing SDS-PAGE (Fig. 3f). SC-specific MAb 9H7 bound only to CH31 SIgA as it contained SC as well as free SC (75-kD protein band) dissociated from CH31 dIgA antibody under nonreducing condition, but MAb 9H7 did not bind to CH31 dIgA that did not contain bound SC (Fig. 3g). SC-specific MAb 9H7 was further tested for detection of SIgA in human BM and plasma as well as in rhesus plasma samples in western blots (Fig. 4). It has been known for several decades that small amounts of SC can appear in the blood circulation.(51) We found that SC MAb 9H7 detected SIgA from both human BM and plasma as protein bands with different molecular weights under nonreducing conditions and also detected SC protein bands dissociated from IgA (Fig. 4b). MAb 9H7 was shown to cross-react with rhesus SIgA from plasma (Fig. 4b).
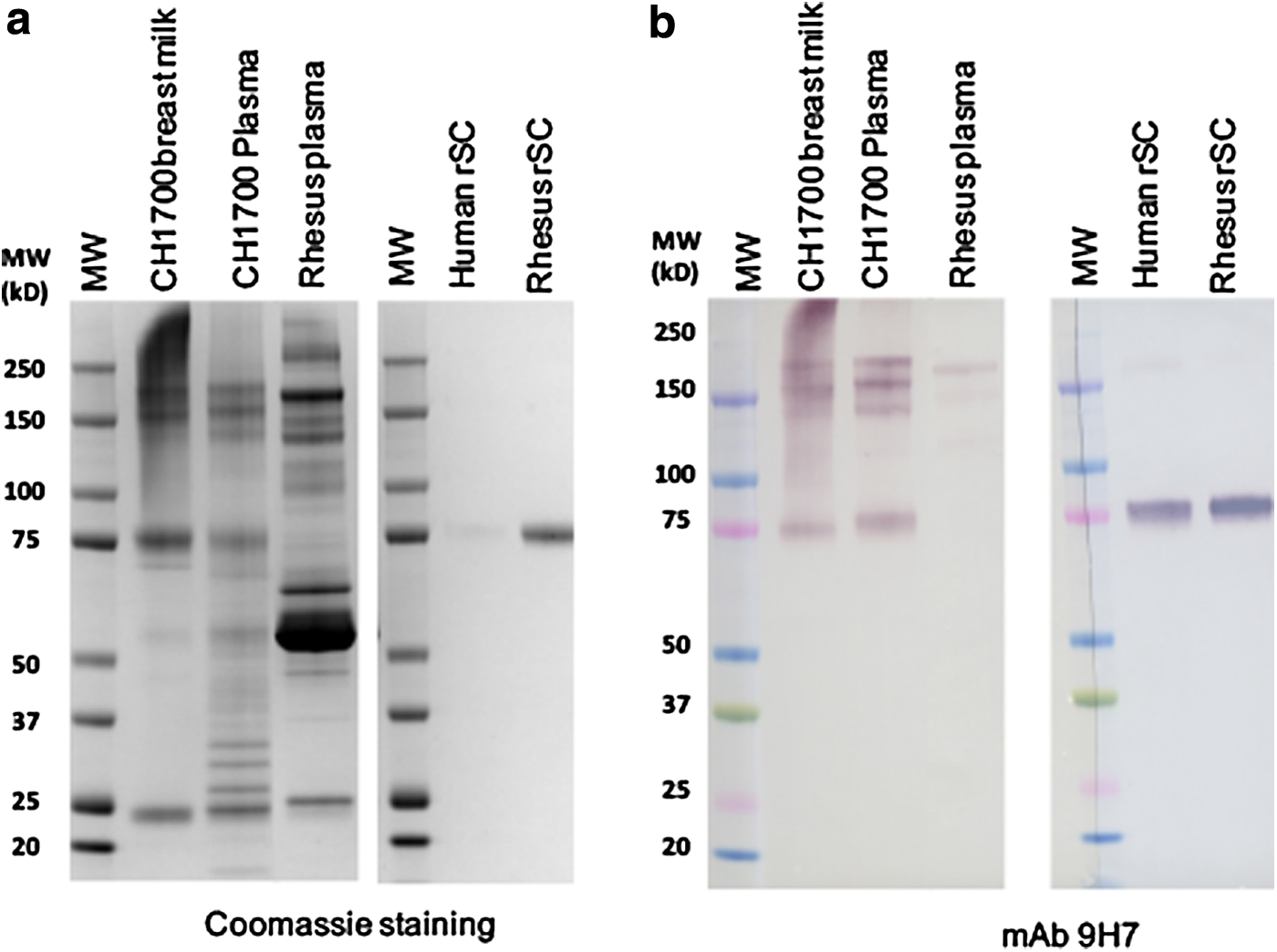
Detection of IgA derived from human BM and human or rhesus plasma. IgA purified from human BM and human (CH1700) and rhesus plasma was fractionated by 4%–20% SDS-PAGE under nonreducing condition. Recombinant human and rhesus free SC was used as control. Gels were stained with Coomassie blue
Immunohistochemistry of human intestine with J-chain and SC MAbs
Both SC MAb 9H7 and J-chain MAb 9A8 were tested by immunostaining on formalin-fixed human tissue sections of histologically normal distal small intestine. MAb 9H7 showed the expected distribution of SC in intestinal crypts, with decreasing intensity of reactivity toward the tips of the villi (Fig. 5A). MAb 9A8 reactivity was compared to that obtained with our rabbit anti-J-chain antibody R-282(46) and was found to produce similar immunostaining patterns in plasmablasts/plasma cells and crypt epithelium (Fig. 5B–E). The epithelial staining was relatively faint compared to that obtained in sections of ethanol-fixed tissue.(52) However, it was specific because its basolateral membrane and cytoplasmic reactivity were as expected, and no FcRn interaction has been observed in previous experiments with human(52) and mouse(21) crypts in ethanol-fixed tissue. In this study, we used formalin-fixed tissue, and despite antigen demasking of cross-linked antigens, we have previously shown that IgA and J-chain are less accessible in the crypt epithelium than in plasmablasts/plasma cells.(53)
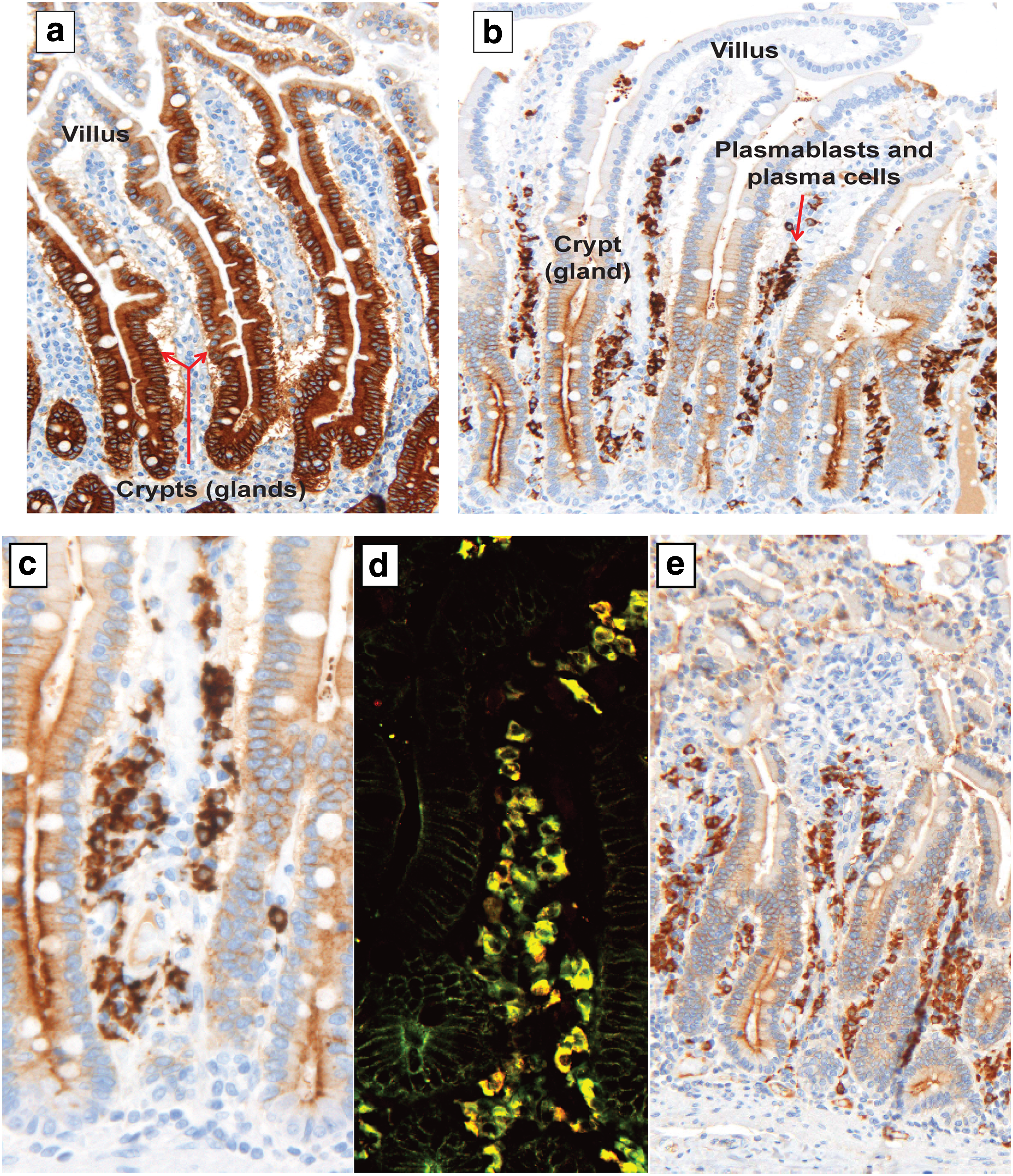
Immunostaining of two key proteins in mucosal immunity, SC and J-chain, on human tissue sections from the distal small intestine.
Discussion
We produced well-characterized MAbs to different forms of IgA and recognized that these reagents can play an important role in understanding the biology of IgA. To characterize different forms of IgA, we produced two mouse MAbs against human J-chain and one MAb against human free SC. They were shown to not only detect recombinant forms of IgA but also recognize dIgA and/or SIgA from human B cell lines and human plasma(51) and BM. Anti-SC MAb 9H7 also cross-reacted with SIgA from rhesus plasma, and we found that human IgA bound to both human and rhesus free rSC. Both J-chain MAb 9A8 and SC MAb 9H7 worked well in immunohistochemistry to identify lamina propria IgA-producing plasmablasts/plasma cells and SIgA-containing epithelial cells in human distal small intestine. The latter immunostaining was, however, relatively faint due to the cross-linking fixative use, namely formalin.(53) Importantly, MAb 9A8 could selectively identify J-chain containing recombinant dIgA versus mIgA.
Because IgA antibodies exist as various forms, including IgA1, IgA2, monomeric, dimeric (polymeric), and SIgAs, we produced recombinant antibodies to these various forms with an HIV-1 bNab (CH31) targeting the HIV-1 gp120 CD4bs(37,38) and a nonneutralizing MAb (7B2) targeting HIV-1 Env gp41 immunodominant region(39) as models. We found that IgA forms of HIV Env antibodies had similar or lower affinity, or weaker activity, than IgG antibodies of the same specificity.(24) However, multivalency of dimeric and larger polymeric Ig forms of IgA and SIgA will enhance their efficacy over monomeric Ig by at least an order of magnitude.(24,54)
Finally, we found that MAb 9H7 could identify SIgA versus dIgA and mIgA molecules. Thus, these new reagents should be of use in the study of the biology of dIgA and SIgA in humans and SIgA in macaques and, as well, in monitoring the production and/or isolation of these forms of IgA.
Footnotes
Acknowledgments
We thank Haiyan Chen, Jamie Pritchett, Sabrina Arora, and Giovanna Hernandez for technical assistance. This work was supported by the Bill and Melinda Gates Foundation (CAVD grant 3830913) and the National Institutes of Health (NIH/NIAID/DAIDS): Center for HIV/AIDS Vaccine Immunology grant (U01 AI067854). P.B. is supported by the Research Council of Norway through its Centres of Excellence funding scheme, project number 179573/40. Authors have no conflict of interest related to this study to declare. Trade names and vendors described were related to the reagents and services used in the study for identification purposes only and do not imply endorsement.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.