
Abstract
CTRP4 (C1q/TNF-related protein 4) is a secreted cytokine homologous to adiponectin, which plays an important role in immunity and metabolism. This study was designed to generate CTRP4 monoclonal antibodies (MAbs). Splenocytes extracted from mice immunized with prokaryotic protein were fused with myeloma cells Sp2/0 to generate hybridoma cells. Three hybridoma strains (16, 33, and 35) were chosen and their MAbs were purified. The specificity and affinity were identified by Western blot and enzyme-linked immunosorbent assay, whereas CTRP4 endogenous expression was identified by immunofluorescence staining. These MAbs could be useful tools for basic research and potential clinical application.
Introduction
C1
Our recent studies document that CTRP4 suppresses colitis and colitis-associated colorectal cancer in mice. In the AOM/DSS mice model, the expression of interleukin-6 as well as the phosphorylation of ERK, STAT3, and Akt in the colon is significantly lower in CTRP4 transgenic mice.(6) Byerly et al. identified that CTRP4 as a unique secreted cytokine modulates food intake and body weight in the hypothalamus.(5) Therefore, CTRP4 may be an important potential indicator in inflammation and metabolism.
Although the functional study of CTRP4 is being examined thoroughly, there is much fundamental information that remains obscure, such as the original source, receptor, and specific subcellular location. CTRP4, just like other members of the CTRP family, is of great importance in immunity and metabolism, as more and more studies have revealed. A number of cytokine-related products have been developed as drugs.(7–9) Knowing more about its fundamental properties may be beneficial to basic study and clinical application.
For further research of the character and function of CTRP4, generation of a CTRP4-specific monoclonal antibody is necessary. Here we report the preparation and identification of a CTRP4 monoclonal antibody.
Materials and Methods
Cell culture
HepG2 cells, 293T cells, and the mouse myeloma SP2/0 cells were cultured in Dulbecco's modified Eagle's medium (GE Healthcare Life Sciences, Logan, UT) added with fetal bovine serum (Life Technologies, Gaithersburg, MD) to a concentration of 10%, 10%, and 15%, respectively. All cells were grown at 37°C in a humidified air incubator with 5% CO2.
Protein expression and purification
The recombinant prokaryotic expression plasmid was cloned by our laboratory. The recombinant plasmids rhCTRP4-pET-32a were transformed into competent Escherichia coli BL21 (DE3) (Biomed, Beijing, China). Competent cells were cultured at 37°C. Then, 0.4 mM isopropyl-β-
Immunization and generation of hybridomas
Beijing Genomics Institute (BGI, Beijing, China) was commissioned to complete the immunization and generation of hybridomas. In summary, BALB/c mice were immunized with a mixture containing purified rhCTRP4 and Freund's complete adjuvant. Two weeks later, the mice were immunized again with adjuvant and recombinant protein. After 2 weeks, a blood sample was extracted from the immunized mice and then tested for titers of the antiserum by enzyme-linked immunosorbent assay (ELISA). The mice with the highest serum titers were chosen for fusion. Their splenocytes were extracted to fuse with mouse myeloma cell line SP2/0. The fused hybridoma cells were seeded individually in 96-well plates. The hybridoma supernatants were tested for titer by ELISA. Forty-one positive hybridoma cells that can recognize rhCTRP4 were chosen for expanded culture.
Preparation and purification of monoclonal antibodies
The hybridoma cells were injected into the peritoneal cavity of 10-week-old female BALB/c mice. The titer of the monoclonal antibodies in the ascites was identified by ELISA. We then harvested all of the ascites and purified them with protein G agarose columns (GE Healthcare Life Sciences, Uppsala, Sweden). The concentration of purified monoclonal antibodies was determined by BCA protein assay.
Antibody isotype determination
Immunoglobulin isotypes of monoclonal antibodies were determined by the mouse monoclonal antibody isotyping kit (Sigma-Aldrich, St Louis, MO).
Western blot analysis
293T cells were transfected with pcDNA3.1-CTRP4-myc-his as an experiment group and the other cells were transfected with empty vector pcDNA3.1-Myc-His (−) B as negative control. Twenty-four hours after transfection, cells were harvested and then lysed in RIPA Lysis Buffer containing 50 mM Tris (pH 7.4), 150 mM NaCl, 1% NP-40, and 0.5% sodium deoxycholate for 30 minutes. The protein concentrations were determined by BCA protein assay according to the manufacturer's instructions. Fifty micrograms of 293T cell lysate was applied to 12.5% SDS-PAGE in SDS sample buffer (62.5 mM Tris-HCl [pH 6.8], 2% SDS, 5% β-ME, 25% glycerol, and 0.02% bromophenol blue). Nitrocellulose membranes (Hybond, ECLTM; Amersham Pharmacia, Little Chalfont, United Kingdom) were used for electrotransfer. After being blocked in 5% BSA in Tris-buffered saline containing 0.1% Tween 20 (TBS-T) for 1 hour at room temperature, the membranes were incubated with culture supernatants or purified monoclonal antibodies in appropriate dilution proportion overnight at 4°C. The horseradish peroxidase (HRP)-conjugated goat antimouse IgG antibody (1:5000 dilution; Cell Signaling Technology, Danver, MA) was incubated for 1 hour at room temperature after the membrane was thoroughly washed. Then we employed the enhanced chemiluminescence Western blot detection substrate (GE Healthcare, Buckinghamshire, United Kingdom) using LAS500 Imager (GE Healthcare, Ontario, CA) to observe the signal followed by washing the membranes in TBS-T.
Enzyme-linked immunosorbent assay
Recombinant protein was diluted by coating buffer (1.59 g Na2CO3 and 2.93 g NaHCO3 in 1 L H2O, pH 9.6) in appropriate proportion. Ninety-six-well plates (Corning, Kennebunk, ME) were coated with the diluted protein 100 μL overnight at 4°C. After being washed twice with PBST (0.05% Tween-20 in PBS) and blocked with 3% BSA for 1 hour at room temperature, each well was incubated with 100 μL diluted antibody or culture supernatants in 3% BSA/PBST for an hour at room temperature. To each well was added 100 μL HRP-conjugated goat antimouse IgG antibodies (1:5000; Abcam Cambridge, United Kingdom) in 3% BSA/PBST for 1 hour followed by washing with PBST three times. Then plates were washed with PBST for five times, after that 50 μL TMB solution was added to each well, and absorbencies were determined after 10 minutes at 450 nm by a microplate reader.
Immunofluorescence staining
HepG2 cells were grown on glass slides. The slides were taken out of culture medium and air dried. Before staining, cells on the slides were fixed in acetone for 5 minutes. After acetone was volatilized, slides were incubated with blocking buffer (5% BSA in PBS) for 1 hour at room temperature. Monoclonal antibodies (diluted 1:100 in 5% BSA/PBS) were dropped to the cells on the slides and incubated for 45 minutes at room temperature. After being gently washed in PBS, cells were stained with FITC-conjugated antimouse IgG (diluted 1:200; Biolegend, San Diego, CA) and were kept out of light for 30 minutes at 4°C. Next, cells were stained with 0.2 μg/mL Hoechst33342 (Sigma-Aldrich, St Louis, MO) in the dark for 3 minutes after being washed three times in precold PBS. After being washed in PBS again, slides were sealed with triglyceride.
Fluorescence microscope observation
HepG2 cells were stained with FITC and Hoechst. After they were sealed in glass slides, the cells were observed under Olympus BX51 fluorescence microscope equipped with an Olympus DP72 camera (Olympus Optical, Tokyo, Japan). For a high quality image, they were observed with Nikon A1 confocal laser microscope (Nikon, Japan).
Results
Generation of hybridoma secreting monoclonal antibodies against CTRP4
The splenocytes, which had the highest antibody titer chosen from an immunized mouse by indirect ELISA, were fused with myeloma SP2/0 cells. Forty-one positive hybridoma clones were achieved and among them 22 isotypes of CTRP4 were identified as IgG.
Reactivity and specificity of purified monoclonal antibodies against rhCTRP4
Vector pcDNA3.1-CTRP4-myc-his was transfected to 293T cells, and 22 positive hybridoma culture supernatants were incubated to overexpression of CTRP4 in the lysate of 239T on Western blot. Then three monoclonal antibodies (MAbs) (MAb 16, MAb 33, and MAb 35) were chosen for purification. The purity of the MAbs was identified by SDS-PAGE (Fig. 1A). Western blot showed that MAb 16, MAb 33, and MAb 35 could react to CTRP4, and the specificity of MAb 33 was better than the others (Fig. 1B).
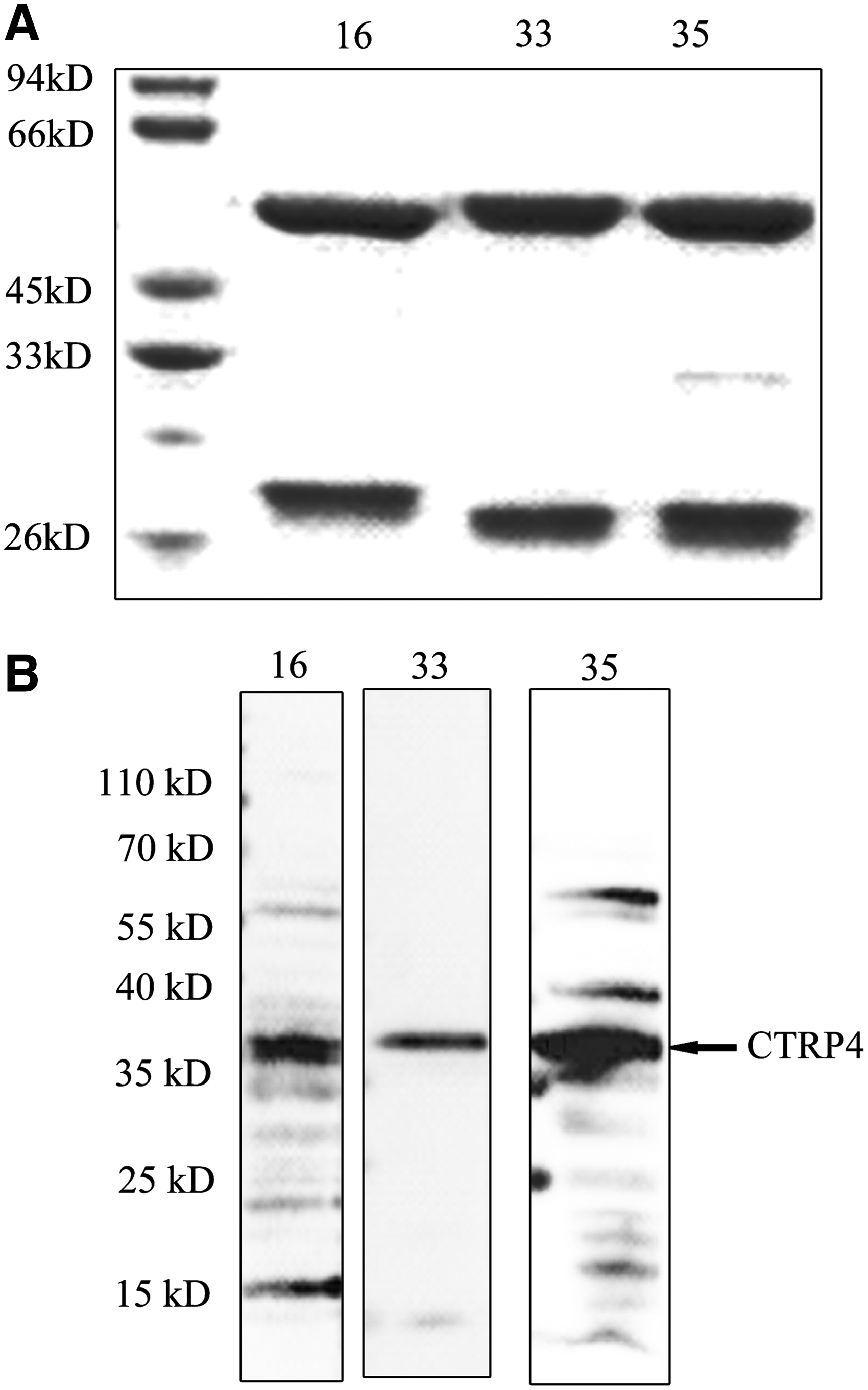
Reactivity of CTRP4 MAbs with overexpressed CTRP4.
Determination of affinity of CTRP4 MAb
The results of the assay suggests that the relative affinity of CTRP4 MAb was high, especially MAb 33 (Fig. 2). CTRP4 MAb could recognize spatial conformation of recombinant eukaryotic CTRP4, and its lower limit is nanogram.

Affinity of anti-CTRP4 MAb analysis by ELISA. Ninety-six-well plates were coated with eukaryotic protein and 3% BSA was used as a control, recognized by three different strains of CTRP4 MAbs:
Subcellular location of CTRP4 MAb in HepG2
CTRP4 has been proven to be a secreted protein, but what the CTRP4 original sources are and how CTRP4 reacts with target cells are still unknown. To help find the answer, we employed immunofluorescence to show the expression of CTRP4 in HepG2 cells. The immunofluorescence scanning showed that MAb 33 was located in the cytoplasm and around the cell nucleus, the recognized site of MAb 35 was in the cytoplasm, and the signal of MAb 16 was weak (Fig. 3). These results indicated that CTRP4 may be located in the cytoplasm and beside the cell nucleus.
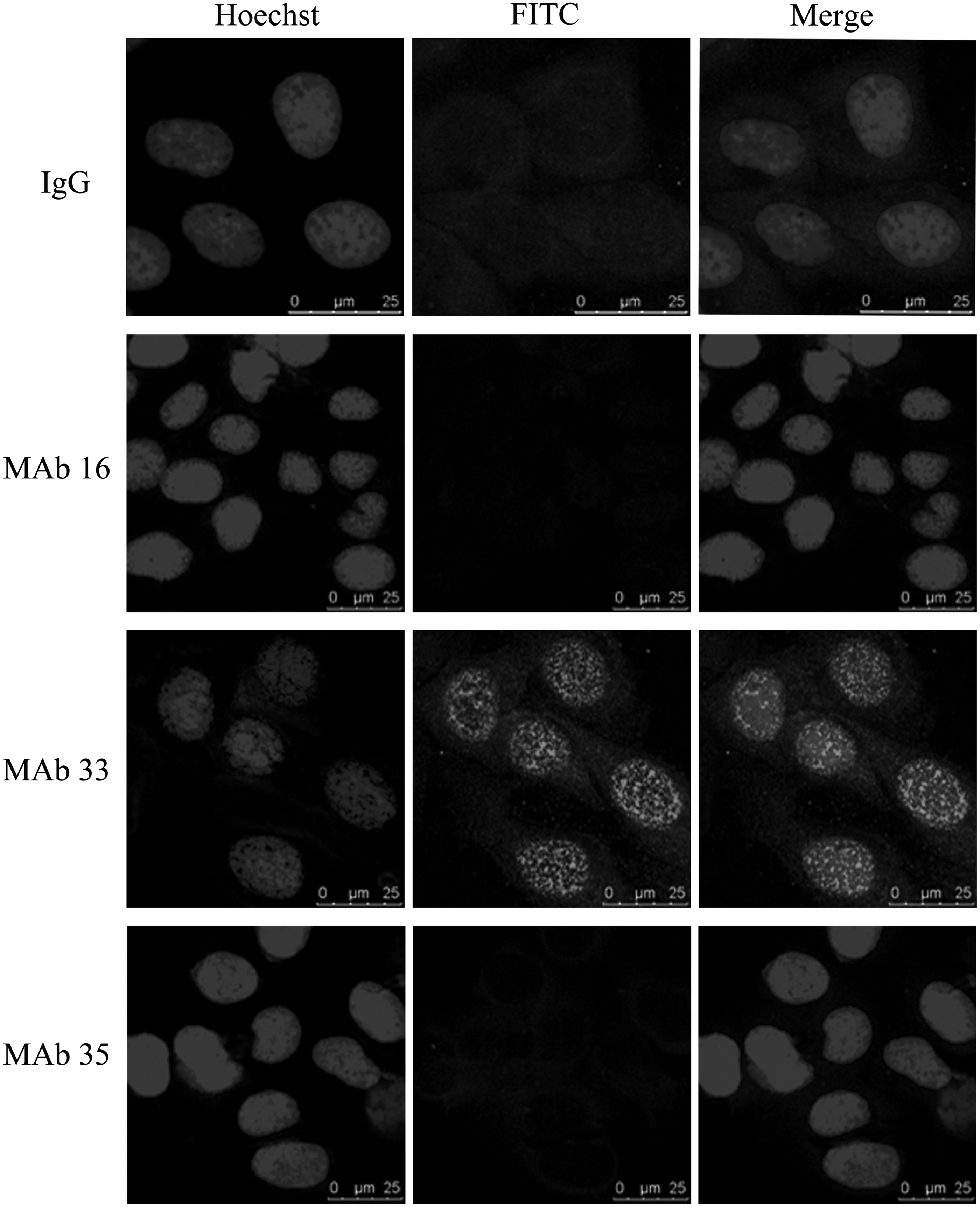
Endogenous expression of CTRP4 in HepG2 cells analyzed by immunocytochemistry. CTRP4 MAbs were used as first antibody, detected by FITC-conjugated antimouse IgG antibody. Normal mouse IgG was used as a negative control.
Discussion
Although CTRP4 has not been studied as extensively as other members of the CTRP family, functional studies of CTRP4 is growing. Therefore, generation of a CTRP4 MAb would help achieve a better understanding of CTRP4. Here, our results suggested that the monoclonal antibodies we generated were relatively specific and high affinity to CTRP4, and CTRP4 may be located in the cytoplasm and beside the cell nucleus.
High efficiency is a characteristic of cytokines,(10) whose biological effects can only be shown through low concentration on blood or other body fluids. An MAb's high affinity to cytokine is of great importance in detection and diagnosis. Our results showed that the MAb was able to recognize CTRP4 in the nanogram range. Studies on CTRP4 indicated that CTRP4 could take effects in this range as well.(4,6) Moreover, mice for monoclonal antibody preparation were immunized by prokaryotic CTRP4, which was not as well folded and assembled as eukaryotic protein. Western blot denaturing the protein spatial conformation cannot eliminate the possibility of recognition of internal epitopes rather than external spatial epitopes. Thus, we proceeded with ELISA detection, coating the plates with eukaryotic protein. Our results showed that CTRP4 MAb could recognize and stick to them. So these MAbs are well qualified for CTRP4 serological test application.
Cytokines are synthesized in the cytoplasm before being secreted out of the cell. However, the original source of CTRP4 is still unclear. The expression of CTRP4 in HepG2 is low, and the three strains of MAbs we purified failed to detect the endogenous CTRP4 by Western blot. Confocal scanning of immunofluorescence showed that CTRP4 is located in the cytoplasm and nucleus. It is the first time that CTRP4 location on cells has been shown. It is hard to tell whether the overlapping epitope with CTRP4 on the nucleus or physiological transduction to the nucleus results in this location. Immunofluorescence indicated that CTRP4 is more likely to locate on the nuclear pore than combine with chromatin. In addition, many cytokines and growth factors or their receptors could be located in the nucleus with the help of the nuclear pore complex.(11–13) The original sources of CTRP4 and distribution still need exploration.
As the functional studies of CTRP4 is increasing, the specific monoclonal antibody we generated in this study could be used as an effective tool for further research on its biological functions, facilitating to complete the fundamental messages of CTRP4, and potentially contributing to basic study and clinical application.
Footnotes
Acknowledgment
This work was supported by grants from the National Natural Science Foundation of China (No. 91129707).
Author Disclosure Statement
No competing financial interests exist.