
Abstract
The interaction between podoplanin (PDPN) and C-type lectin-like receptor 2 (CLEC-2) is involved in tumor malignancy. We have established many monoclonal antibodies (mAbs) against human podoplanin using the cancer-specific mAb (CasMab) technology. LpMab-21, one of the mouse antipodoplanin mAbs, is of the IgG2a subclass, and its minimum epitope was determined to be Thr76–Arg79 of the human podoplanin. Importantly, sialic acid is linked to Thr76; therefore, LpMab-21 is an antiglycopeptide mAb (GpMab). In this study, we investigated whether LpMab-21 shows antibody-dependent cellular cytotoxicity (ADCC) and complement-dependent cytotoxicity (CDC) against human podoplanin-expressing cancer cell lines in vitro and also studied its antitumor activities using a xenograft model. LpMab-21 showed high ADCC and CDC activities against not only podoplanin-expressing Chinese hamster ovary cells but also LN319 glioblastoma cells and PC-10 lung cancer cells, both of which endogenously express podoplanin. Furthermore, LpMab-21 decreased tumor growth in vivo, indicating that LpMab-21 could be useful for antibody therapy against human podoplanin-expressing cancers.
Introduction
P
Although many antihuman podoplanin monoclonal antibodies (mAbs) are commercially available, almost all the mAbs react with the N-terminus of human podoplanin.(6,25–29) In contrast, we have used the cancer-specific mAb (CasMab) technology to produce antiglycopeptide mAbs (GpMabs) against human podoplanin.(30–39) Recently, we have successfully developed a novel antihuman podoplanin mAb, LpMab-21, which recognizes a sialylated glycopeptide epitope.((40) LpMab-21 is one of the GpMabs, but not a CasMab. Importantly, LpMab-21 is the first mouse antihuman podoplanin mAb of the IgG2a subclass, which was generated using the CasMab technology.(30–36,38,39)
Results and Discussion
We have produced several antihuman podoplanin mAbs using the CasMab technology; however, the isotypes of the mAbs are IgG1 (seven clones) and IgG3 (one clone), which do not induce antibody-dependent cellular cytotoxicity (ADCC) or complement-dependent cytotoxicity (CDC). The applications of mouse IgG3 mAbs are limited because they often aggregate.((41) In this case, we have had to convert them to human IgG1 to investigate whether these mAbs cause ADCC and CDC.((33) In contrast, the mouse IgG2a subclass can induce ADCC and CDC; therefore, we investigated whether LpMab-21 can induce ADCC and CDC against human podoplanin-expressing cancer cell lines. As shown in Figure 1A, LpMab-21 demonstrated ADCC against LN319, the Chinese hamster ovary (CHO)/human podoplanin, and PC-10 cell lines, whereas it did not induce ADCC against human podoplanin-negative parental CHO cells. Similarly, LpMab-21 induced CDC against LN319 and PC-10 cell lines (Fig. 1B). We previously demonstrated that LN319 expresses human podoplanin at a higher level than does PC-10((30); therefore, ADCC and CDC might depend on the human podoplanin expression levels in these cell lines.
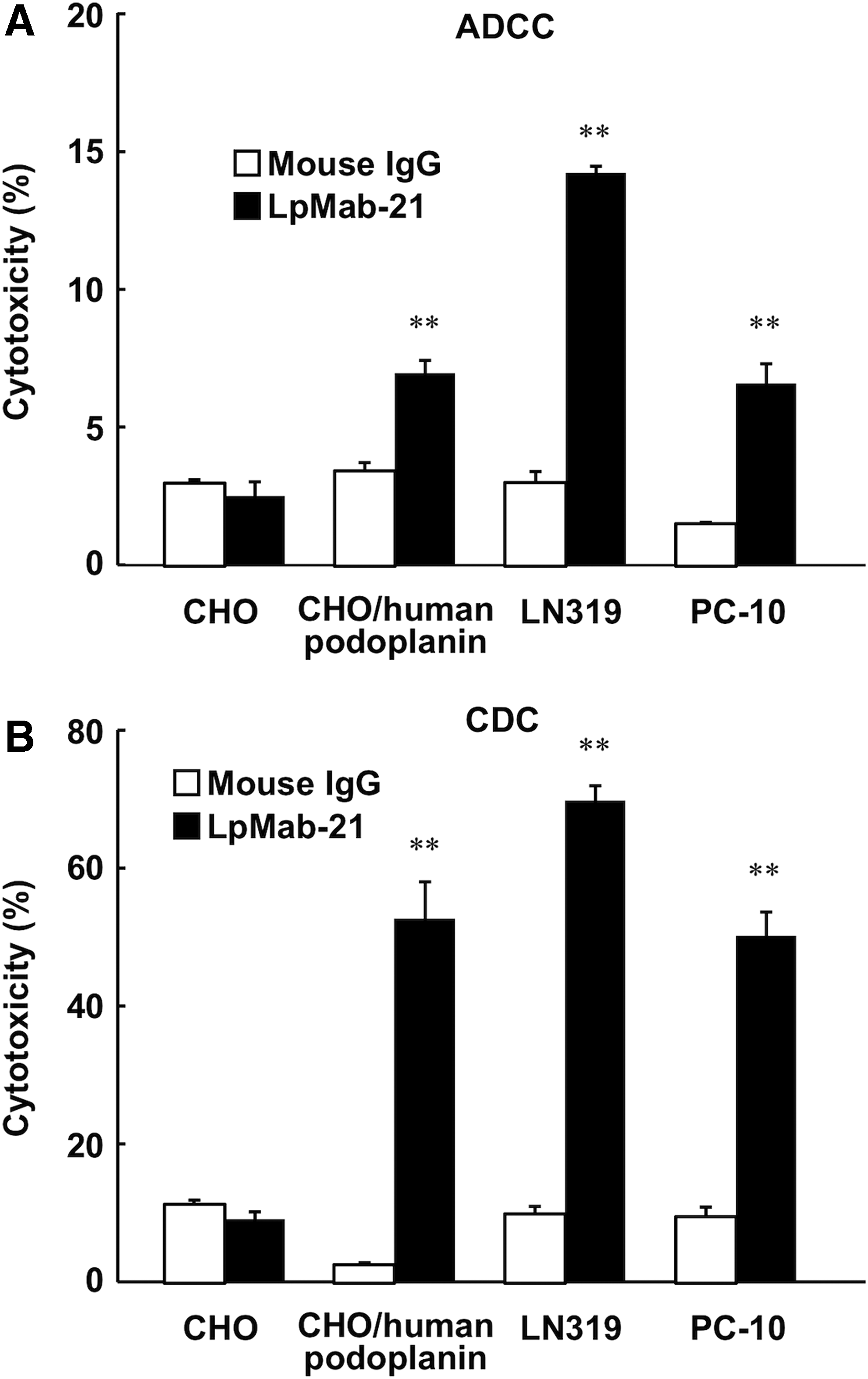
Antibody-dependent cellular cytotoxicity (ADCC) and complement-dependent cytotoxicity (CDC) of LpMab-21 against human podoplanin-expressing cell lines.
To investigate the antitumor activity of LpMab-21 on primary tumor growth in vivo, CHO/human podoplanin cells were subcutaneously implanted into flanks of nude mice. LpMab-21 or control mouse IgG (clone PMab-2)((42) was injected into the peritoneal cavity of the mice once weekly for 4 weeks (n = 6 each). PMab-2 was raised against rat podoplanin and was shown to not cross-react with human podoplanin.((42) Tumor formation was observed in five mice from the control group (tumor incidence on day 35: 83.3%, 5/6; Supplementary Fig. S1). In contrast, LpMab-21 dramatically reduced the tumor development (tumor incidence on day 35: 50%, 3/6; Supplementary Fig. S1). The tumor volume was significantly reduced by LpMab-21 treatment (Fig. 2). These results indicate that administration of LpMab-21 inhibited the primary tumor growth of CHO/human podoplanin cells. In our previous study, only CDC of a human–mouse chimeric anti-PDPN mAb could show antitumor activity because human NK cell was not added in this xenograft model.((43) In contrast, we also showed that ADCC of a human–mouse chimeric anti-PDPN mAb is more important than CDC in another study.((9) Therefore, we think that both ADCC and CDC are important to induce antitumor activity in the xenograft model.
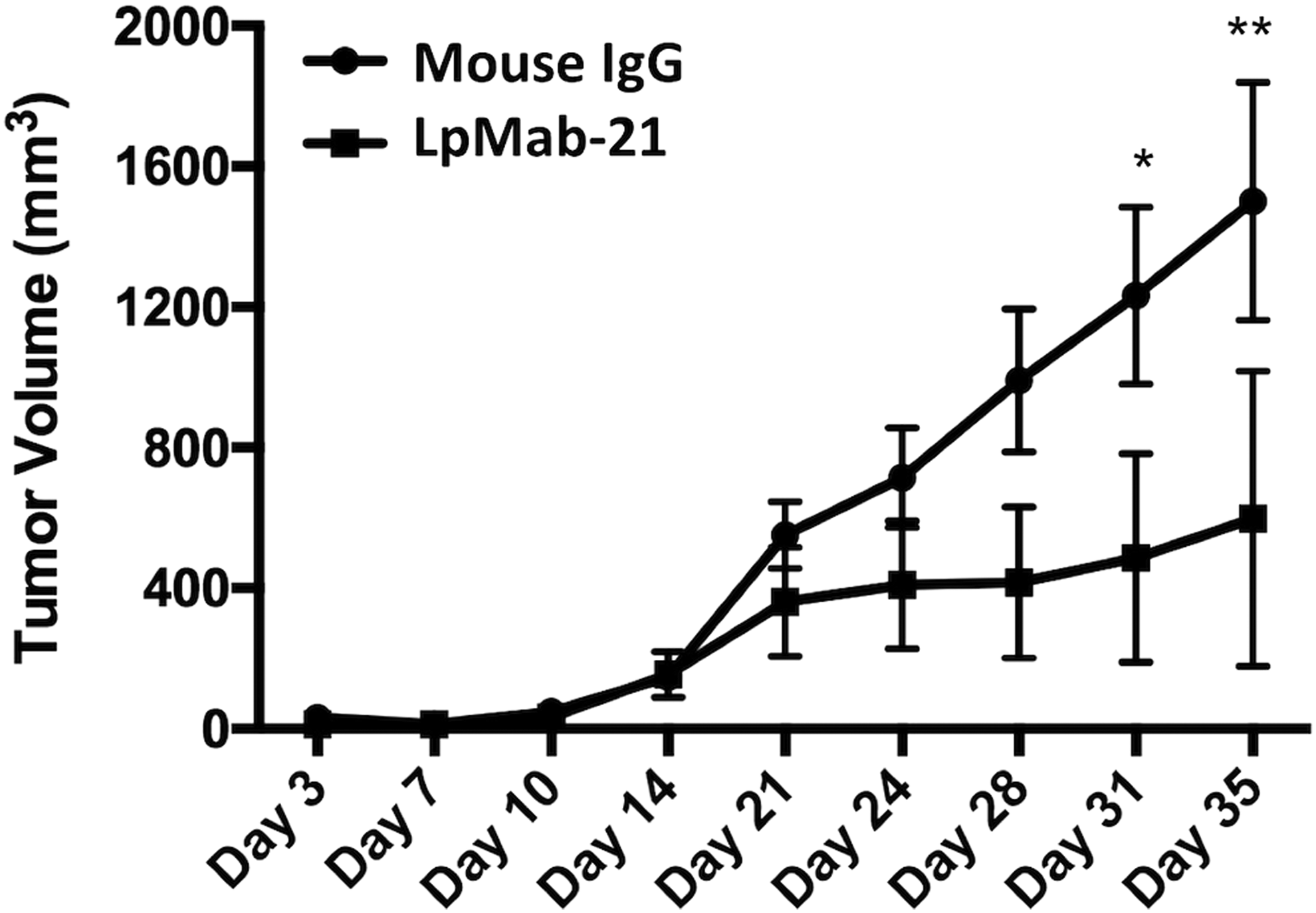
Antitumor effects of LpMab-21 on primary tumor development. CHO/human podoplanin cells (3 × 106 cells/100 μL) were subcutaneously inoculated into BALB/c nude mice. After 1 day, 100 μg of LpMab-21 or control mouse IgG (clone PMab-2) was injected into the peritoneal cavity of the mice. The antibodies were injected once weekly for 4 weeks (control group: n = 6; LpMab-21 group: n = 6). The tumor diameter was measured at intervals of 3 to 4 days and was calculated using the following formula: tumor volume = W2 × L/2, where W is short diameter and L is long diameter. *p < 0.05; **p < 0.01 with two-way analysis of variance.
Taken together, LpMab-21 could be useful for antibody therapy against human podoplanin-expressing cancers. Our developed CasMabs as well as LpMab-21 could be applied to the novel antitumor reagents, including T cells and viruses,((44) to give strict specificity against tumor cells.
Footnotes
Acknowledgments
This work was supported, in part, by project for utilizing glycans in the development of innovative drug discovery technologies from Japan Agency for Medical Research and development, AMED (Y.K.), by the Basic Science and Platform Technology Program for Innovative Biological Medicine from AMED (Y.K.), by the Platform for Drug Discovery, Informatics, and Structural Life Science (PDIS) from AMED (Y.K., T.M.), by Translational Research Network Program from AMED (Y.K.), by JSPS KAKENHI Grant Number 26440019 (M.K.K.) and 16K10748 (Y.K.), and by the Regional Innovation Strategy Support Program from the Ministry of Education, Culture, Sports, Science and Technology (MEXT) of Japan (Y.K.). This work was performed, in part, under the Cooperative Research Program of Institute for Protein Research, Osaka University, CR-16–05, and by the Grant for Joint Research Project of the Institute of Medical Science, the University of Tokyo. The authors would like to thank Enago (
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.