
Abstract
A monoclonal antibody (Mab) against lycopene was developed from hybridoma clones obtained from BALB/c mice immunized with trans-isomer of lycopene (t-lycopene, t-LC) conjugated with colloidal gold particles. An alternating immunization schedule which included injection of both formulations of immunogen (without and with Freund's adjuvant) was most effective in the elucidation of a measurable immune response to the t-Lycopene conjugate. Selected hybridoma clones were able to produce an Mab positive in competition assay. In particular, preincubation of 6B9 Mabs with t-LC abolished the ability of 6B9 Mabs to bind LC in the competition assay. Mabs produced by other clones (4F10, 4A3, and 3B12) worked similarly. Analysis of antigen specificity showed that 6B9 Mab raised against t-LC did not recognize other carotenoids such as lutein and carotene. Mab 6B9 was shown to recognize lycopene on a glass surface and in the settings of indirect immunofluorescence experiments performed in cultured hepatocytes and alveolar macrophages incubated with and without lycopene, as well as in sebum and corneocyte specimens from the skin of volunteers supplemented with nutraceutical formulation of lycopene. Newly generated Mabs against lycopene may provide a valuable tool for different analytical assays of lycopene content in various biological, agricultural, and food products.
Introduction
L
Timely identification of lycopene deficiencies and their effective correction with dedicated food or nutraceutical supplementation would be significantly improved and have higher compliance if both health practitioners and patients could have more accessible laboratory or even point-of-care quantitative test(s) for lycopene measurement.
High-performance liquid chromatography along with other analytical procedures remains the preserve of specialized laboratories and is the only option for quantification of lycopene in different biological materials.
In the present study, we report on the development of new monoclonal antibodies against lycopene, which can be used for evaluation of lycopene content in different biological materials, including agricultural, food, and clinical specimens.
Materials and Methods
All trans-Lycopene (t-LC), lutein, carotene, HAuCl4 bovine serum albumin (BSA), Freund's adjuvant, Liquid Substrate System for enzyme-linked immunosorbent assay (ELISA) reagents and other chemicals, solvents, and supplies were purchased from Sigma-Aldrich (St. Louis, MO). ELISA plates were purchased from Greiner Bio-One (No. 762070; Frickenhausen, Germany). Female BALB/c mice aged 3 months were ordered from the Puschino Breeding Facility (Moscow, Russia). All aqueous solutions, including high-pressure liquid chromatography (HPLC) buffers, were prepared using Millipore Milli-Q grade water.
Lycopene-gold conjugate
Lycopene-gold conjugate was used for the immunization protocol. Colloidal gold nanoparticles (GNPs) with a size of 15 or 30 nm (GNPs15 and GNPs30) were prepared as previously described(8,9)
In brief, 0.5 mL of 1% HAuCl4 was added to 48.0 mL of boiling water and stirred for 2 minutes. Addition of a 1% solution of sodium citrate was made immediately. To achieve the formation of GNPs with a size of 15 nm, 1.5 mL of sodium citrate was added. GNPs with a size of 30 nm were obtained by addition of 0.72 mL of sodium citrate. The reaction mixture was boiled for 20 minutes and cooled down to room temperature.
Lycopene was dissolved in EtOH (0.5 mg/mL), and the solution was heated to 80°C until the crystals were completely dissolved. The GNPs solution was mixed with lycopene-EtOH at a ratio 5:1 (v/v) and kept at room temperature for 20 minutes. The conjugated obtained was immediately used for immunization injections.
Antibody generation
All animal procedures were described in the study protocol approved by the local Animal Research Ethical Committee at the Moscow Institute of Virology. Suspensions of t-LC- GNPs conjugates were freshly prepared, either mixed, or not mixed, with the complete Freund adjuvant and injected intraperitoneally. Mice were immunized five times with a 2-week interval between injections of conjugates.
Serum evaluation
Serum specimens from the immunized mice were routinely screened using indirect ELISA after the second round of immunization.
Titer check
Each well of ELISA microplates (Greiner Bio-One 96-well) was coated with 100 μL of lycopene diluted in absolute ethanol (10 μg/mL) and then dried out at room temperature.
Control sera from nonimmunized mice and sera from immunized mice were diluted with PBST supplemented with 1 mg/mL BSA. One hundred milliliters serial dilutions ranging from 1:1000 to 1:128,000 were pipetted into the wells of the ELISA plates, with further incubation of the plates at 37°C for 1 hour. After three washes with PBST, addition of peroxidase-labeled goat antibody against mouse IgG was made. Further incubation of the plates at 37°C for 1 hour was followed by three PBST washes and addition of 100 μL/well of tetramethylbenzidine chromogenic substrate. After 30 minutes incubation at room temperature, the color reaction was stopped by adding 50 μL/well 1 N H2SO4. The plates were read at 450 nm (Multiscan EX; Thermo Scientific, Waltham, MA). The titers were measured for each individual mouse after each round of the immunization cycle.
Hybridoma construction
After 72 hours following the final intravenous boost, the spleens of anesthetized mice (positive responders) were removed and dispersed, and the resulting splenocytes were fused with Sp-2 cells and incubated in HAT medium according to the conventional protocol.(9) Positive wells were inspected microscopically for cluster formation, and the supernatants were tested for antibody presence by indirect ELISA. Positive clones were subcloned by limiting dilution protocol using spleen feeders. Antibodies were isotyped with a Mouse Monoclonal Antibody Isotyping Reagent (Sigma-Aldrich). The hybridoma cell line was submitted and deposited at the Institute of Genetics and Selection of Genetics and Selection of Industrial Microorganisms (Submission Inquiry No. 194/16).
t-LC with colloidal gold was used as a coating agent for an indirect competitive ELISA as described above. Serum specimens obtained from immunized mice were used as a source of primary antibody. Increasing concentrations of nonconjugated t-LC mixed with aliquots of sera or ascitic fluid were preincubated on ice for 1 hour before inoculation into the wells of the ELISA microplate.
Validation of t-LC-specific antibody
Validation of t-LC antibody was also performed by immunofluorescence analysis in cultured cells incubated with oil-formulated lycopene. Stock oil solutions of lycopene (15%) were purchased from LycoRed (Basel, Switzerland) and kept at–20°C. For studies in cultured cells, the stock solution was dissolved in DMSO at concentrations of 0.75, 1.5, and 3.0 μg/mL. C B10.MLM, a cell line of alveolar macrophages, was obtained from Prof. AS Apt (Institute of Tuberculosis, Moscow, Russian Federation). Cells were grown in 5% CO2 in DMEM supplemented with 2 mM glutamine and 10% FCS. B10.MLM cells grown on coverslips were incubated with lycopene for 46 hours. Cells were then washed with PBS twice, fixed with 3% formaldehyde/0.025% glutaraldehyde at room temperature for 20 minutes, and permeabilized with 0,3% saponin. After blocking with 3% BSA for 30 minutes, cells were stained with the primary anti-lycopene antibodies (6B9) and FITC-conjugated anti-mouse IgM (μ-chain specific-FITC). Cells were visualized using a Nikon Eclipse 50i fluorescence microscope at × 1000 magnification.
Results
Lycopene immunizations carried out with complete and incomplete Freund's adjuvant mixed with t-LC–GNPs15 (first and second groups, Table 1) were accompanied by the appearance of LC-specific antibodies in serum beginning the second week after immunization in the range of 1:800 to 1:1000. However, after the third immunizing injection, mice from groups 1 and 2 became sick and died, suggesting a toxic effect from GNPs-conjugated lycopene injected with Freund's adjuvant.
cAF, complete Freund's adjuvant; GNPs, gold nanoparticles; iAF, incomplete Freund's adjuvant.
Very low antibody response was seen in the mice injected with t-LC–GNPs15и t-LC–GNPs30 without adjuvant (Groups 3 and 4, Table 1). Much better immune response was seen in the group 5 of mice injected with lower amount of immunogen (17 μg/mouse) and reduced number of immunizing injections (three injections instead of five) and further buster injection of unconjugated t-LC.
One of the immunized mice (No. 3) had a distinctive antibody titer (1:1600) and was used for the fusion procedure (Fig. 1). Therefore, the immunization protocol used was considered to be a most effective way to achieve a measurable immune response to t-LC.
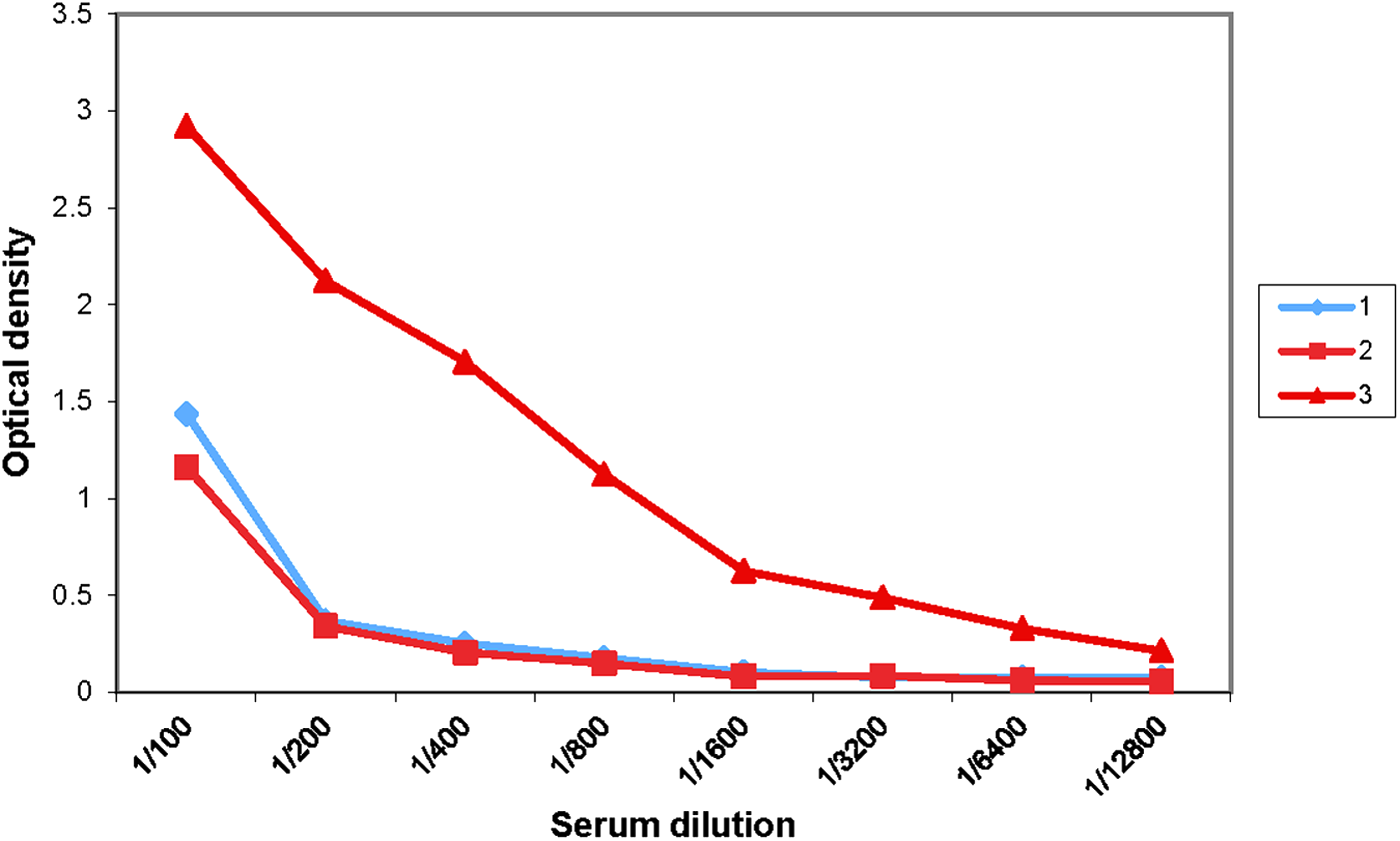
Antibody titer immunized mice belonging to the fifth immunization group.
Fusion protocol allowed us to obtain over 500 primary hybridoma clones selected in the HAT medium. Screening of culture medium for the presence of t-LC-specific antibodies was conducted from day 8 of clone cultivation and lead to the establishment of 24 hybridoma clones producing antibody specifically recognizing t-LC. After subcloning, four hybridoma cell lines producing LC-specific IgM antibodies were established. Clone characteristics are shown in Table 2.
All clones mentioned above had some differences in ability to produce Mab (Fig. 2).

Comparison of ascitic fluids from different clones in enzyme-linked immunosorbent assay (ELISA).
Analysis of antigen specificity showed that 6B9 Mab raised against t-LC did not recognize other carotenoids such as lutein and carotene (Fig. 3A, B).
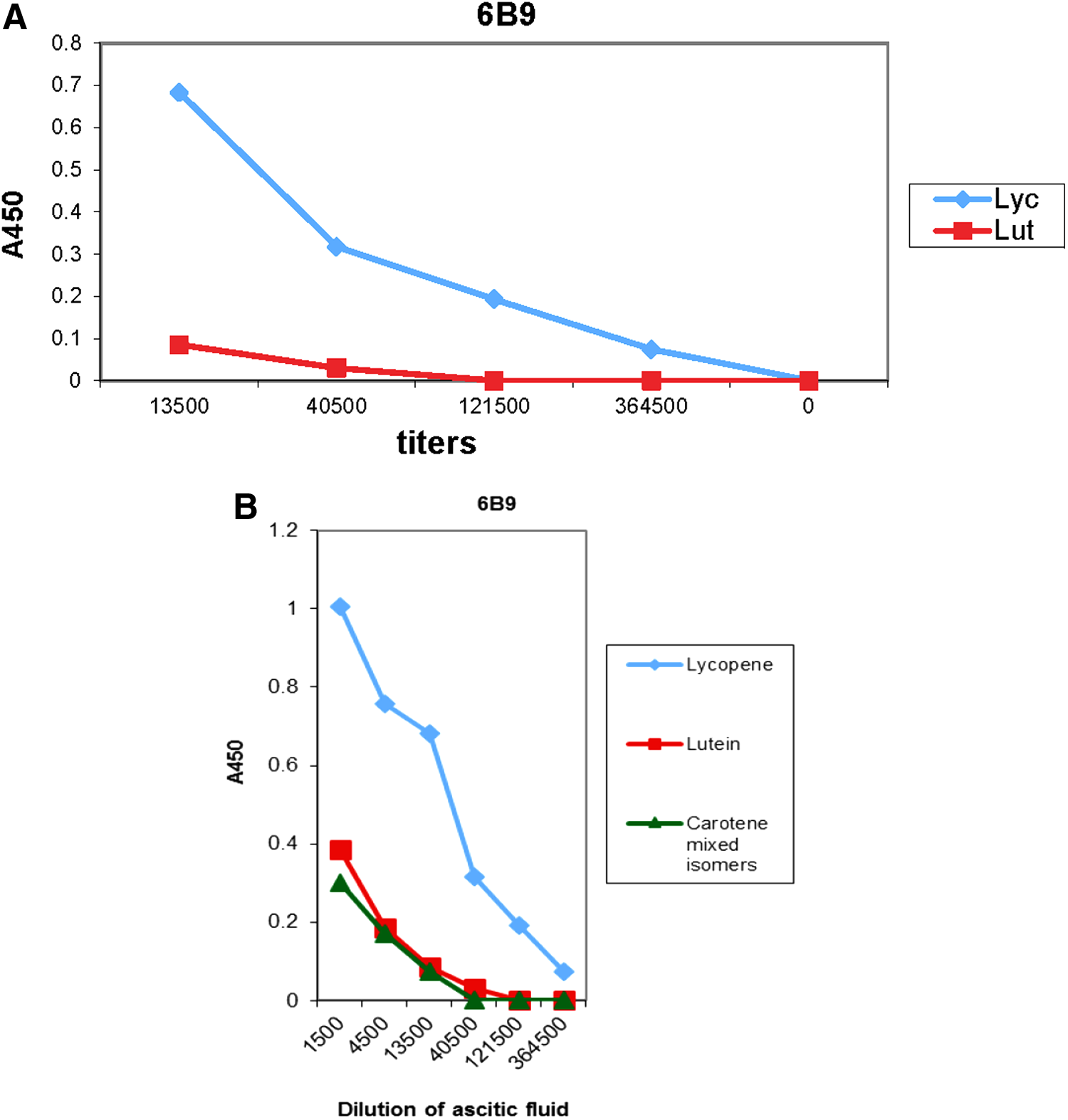
Recognition of lycopene, lutein, and carotene in ELISA by Mab 6B9.
The newly developed Mab 6B9 against t-LC was validated in different systems.
First of all, clones were capable of t-LC detection in the indirect immunoassay performed on a glass slide surface (Fig. 4A). Second, Mab 6B9 was used in immunohistochemistry analysis. It was possible not only to detect t-LC but also to visualize its intracellular distribution within cultured human hepatocytes and alveolar macrophages, following their incubation with an oil solution of t-LC for 48 hours (Fig. 4B).
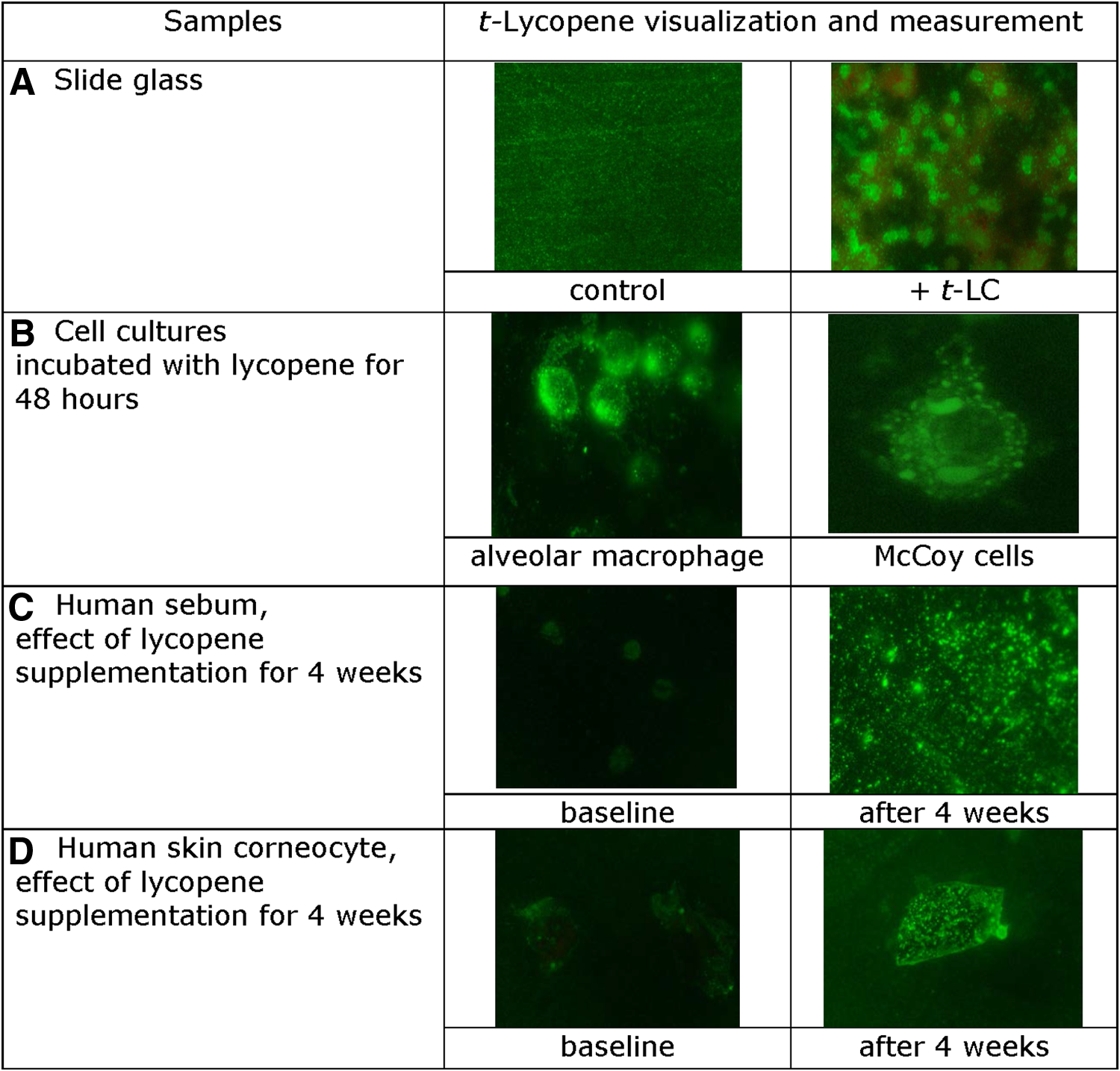
Immunofluorescent detection of t-Lycopone.
Figure 4C and D show that it was possible to measure changes in t-LC presence in sebum and corneocytes in skin imprints of volunteers, after 4 weeks of oral supplementation with 7 mg lycopene supplement (Lycotec, United Kingdom).
Discussion
Immunization protocols based on the use of colloidal gold particles open new possibilities for the generation of highly specific antibodies against various substances as well as for immunodiagnostics and immunotherapy. As recently reported,(10) injections of antigens or haptens with colloidal gold particles promote their delivery to dendritic cells and enhance lymphocyte proliferation and antibody response at much greater levels than other standard immunization techniques. Colloidal particles allow the generation of new antibodies against substances whose immunogenic properties were questioned for many decades.
In the present study, we report on the development of new monoclonal antibodies capable of lycopene recognition, which were raised by immunization of mice with t-lycopene associated with colloidal gold particles. To the best of our knowledge, this is the first description of the successful raising of monoclonal antibodies against a member of carotenoid family. As we have shown above, these antibodies bind specifically to lycopene in the ELISA test. All four hybridoma clones established synthesize Mabs, which recognize lycopene, did not bind to another major carotenoid lutein, which has similar antioxidant properties and structure.(11) Moreover, Mab 6B9 has a unique ability to recognize lycopene inclusions in the settings of indirect immunofluorescence experiments.
In particular, it was shown that Mab 6B9 could not only detect but also visualize lycopene distribution within cultured McCoy cells and alveolar macrophages, which were preincubated with this carotenoid for 48 hours. These results, to the best of our knowledge, for the first time, demonstrate intracellular localization of lycopene in intact cells, predominantly in their cytoplasm.
Moreover, these antibodies were able to recognize and measure changes in t-LC concentration in sebum and corneocytes from the skin of volunteers(12) after supplementation with lycopene nutraceutical for 4 weeks.
However, there is a theoretical challenge to explaining the mechanism underlying antibody response to such a low-molecular weight substance as lycopene. Lycopene is a hydrocarbon compound of relatively low molecular weight (536.89 g·mol−1) belonging to the tetraterpene group. It has 11 conjugated double bonds mediating its antioxidant activities.(1) Lycopene is one of the most powerful antioxidants whose antiradical activity exceeds the antioxidant potential of tocopherol, beta-carotene, and ascorbic acid.(13) The highly unsaturated chemical structure of lycopene predetermines its ability to react with many substances containing or generating singlet molecular oxygen such as peroxyl radicals, hydrogen peroxide, and hypochlorite.(1,13) Therefore, molecular associations between lycopene and oxidized proteins or carbohydrates may confer on the lycopene molecule hapten properties capable of elucidating the immune response. Colloidal gold may promote such temporary associations of lycopene with macromolecules required for the initiation of immune response. Nevertheless, the mechanism of antibody response to lycopene administration with colloidal gold particles remains unclear and should be further evaluated.
It should be stated also that the development of lycopene-specific Mab opens up a number of exciting new possibilities. This includes the ability to detect lycopene and assess its intracellular distribution and distribution in tissues in vitro, ex vivo, and in situ, without affecting integrity of the cell or tissue structures, both in an experimental setting or for diagnostic purposes.
In addition, these antibodies could be used for the development of assays of different design not only for laboratory applications but also for point-of-care self-monitoring tests. The cost and relative simplicity of these tests would make screening and detection of lycopene deficiencies, and also their correction by changes in diet or nutraceutical supplementation, significantly more efficient.
At present, the only method of choice for lycopene quantification in biological or clinical specimens is HPLC, which is expensive and requires specialized chemistry laboratory expertise. Development of new analytical and diagnostic tools based on newly established lycopene-specific Mab may allow highly efficient and simple protocols for quantification of lycopene in biological specimens, transforming lycopene quantification procedure into a routine laboratory method.
Moreover, use of these antibodies as the basis for new point-of-care tests could make prevention and correction of lycopene deficiencies more effective, and therefore be an important tool for personalized nutrition in support of the cardiovascular system, cancer prevention, and healthy ageing.
Footnotes
Acknowledgment
The authors express appreciation to Dr. Vladimir A. Bogaturev for sharing valuable reagents.
Author Disclosure Statement
No competing financial interests exist.