
Abstract
IgA nephropathy (IgAN) is a globally common primary glomerulonephritis characterized by an elevated level of serum IgA and immune complex deposition in the mesangial area. In the serum of patients with IgAN, the hinge region of IgA1 immunoglobulin contains aberrantly glycosylated O-glycans deficient in galactose, which is normally added to the core 1 O-glycan structure by core 1 synthase, glycoprotein-N-acetylgalactosamine 3-beta-galactosyltransferase 1 (C1GALT1), the key enzyme in the process of glycosylation. It is unknown if single-nucleotide polymorphisms rs1047763 and rs1008898 of C1GALT1 increase the risk of IgAN. We enrolled 5 subjects in this meta-analysis, including a total of 1693 IgAN patients and 1864 control subjects. We performed meta-analysis on associations between rs1047763, rs1008898, and IgAN using the allele model, dominant model, recessive model, and additive model. We found that there was no relationship between rs1047763 and rs1008898 in C1GALT1 and susceptibility to IgAN.
Introduction
I
A major characteristic of IgAN is IgA deposition (mainly the IgA1 subclass) in mesangial cells.(9,10) The hinge region of IgA1 immunoglobulin contains O-linked glycans with N-acetylgalactosamine (GalNAc), galactose (Gal), and sialic acid. The linkage of Gal to GalNAc is catalyzed by enzyme core 1 synthase, glycoprotein-N-acetylgalactosamine 3-beta-galactosyl transferase 1(C1GALT1). A final extension of the core 1 glycan structure by sialic acid added to GalNAc or Gal is the normal pattern of IgA1 glycosylation.(11,12) However, serum IgA1 in IgAN patients lack Gal in one or more of the hinge region O-linked glycans, thus presenting with a terminal GalNAc or sialylated GalNAc (Gal-deficient IgA1 or Gd-IgA1),(13) which is recognized by autoantibodies in susceptible individuals and results in immune complex formation enriched in Gd-IgA1.
The C1GALT1 gene is located in 7p21.3 and encodes the protein responsible for generating the common core 1 O-glycan structure. A study has shown that this gene plays an important role in the glycosylation of IgA1 molecules.(14) Xia and McEver(15) showed that C1GALT1 is a key regulatory gene in the process of glycosylation, but the relationship between single-nucleotide polymorphisms (SNPs) in the C1GALT1 gene and IgAN is not clear. Bertinetto et al.(16) demonstrated that the rs1008898 and rs7790522 SNPs of C1GALT1were associated with IgAN. Furthermore, a study by Wang et al.(6) examining IgAN patients in southern China reported that rs1008898 of C1GALT1 and rs340883 of ILRA31 interacted with each other and were associated with IgAN, but the occurrence of either SNP alone was unrelated to IgAN. Therefore, the association of a particular SNP of the C1GALT1 gene with IgAN has not been shown. Pirulli et al.(17) showed that the rs1047763 allele G and the GG genotype were significantly more frequent in IgAN patients than in controls. However, results from studies by Mostowska et al.,(18) Li and Lu,(19) and Li et al.(20) have been controversial. Since there has been no previous meta-analysis examining the association between C1GALT1 SNPs and IgAN risk, we conducted this meta-analysis to provide a more precise estimation of the relationship between them.
Materials and Methods
Our search strategy to examine the association of rs1008898 and rs1047763 C1GALT1 gene polymorphisms with the risk of IgAN was as follows. We searched the electronic databases of PubMed, EMBASE, Wanfang, and Weipu before October 15, 2015. The searching words were “(IgA nephropathy OR IgAN) AND (polymorphism OR polymorphisms OR variant OR variants) AND (C1GALT1).”
Inclusion and exclusion criteria
Inclusion criteria
(1) Cases were IgAN patients, (2) the diagnostic method should be kidney biopsy, (3) studies were case–control studies, (4) studies evaluated the association of rs1008898 or rs1047763 with IgAN risk, and (5) articles provided data about C1GALT1 genotype distributions.
Exclusion criteria
(1) Review articles, case reports, and editorials, (2) patient groups without IgAN, (3) results not pertinent to C1GALT1 rs1008898 or rs1047763 polymorphisms, (4) investigations of the role of C1GALT1 gene in the pathogenesis of disease and the clinical manifestations of disease, and (5) failure to use healthy populations as controls. When there were multiple publications of the same data from the same group, only the study with the largest sample size was included.
We finally enrolled four reports(17–20) examining rs1047763 in 1165 IgAN patients and 1316 control subjects, and 3 studies(6,19,20) examining rs1008898 in 1377 IgAN patients and 1217 control subjects. Two of the studies(19,20) examined both SNPs in the same groups, so a total of five independent studies examining either SNP were enrolled (Fig. 1).
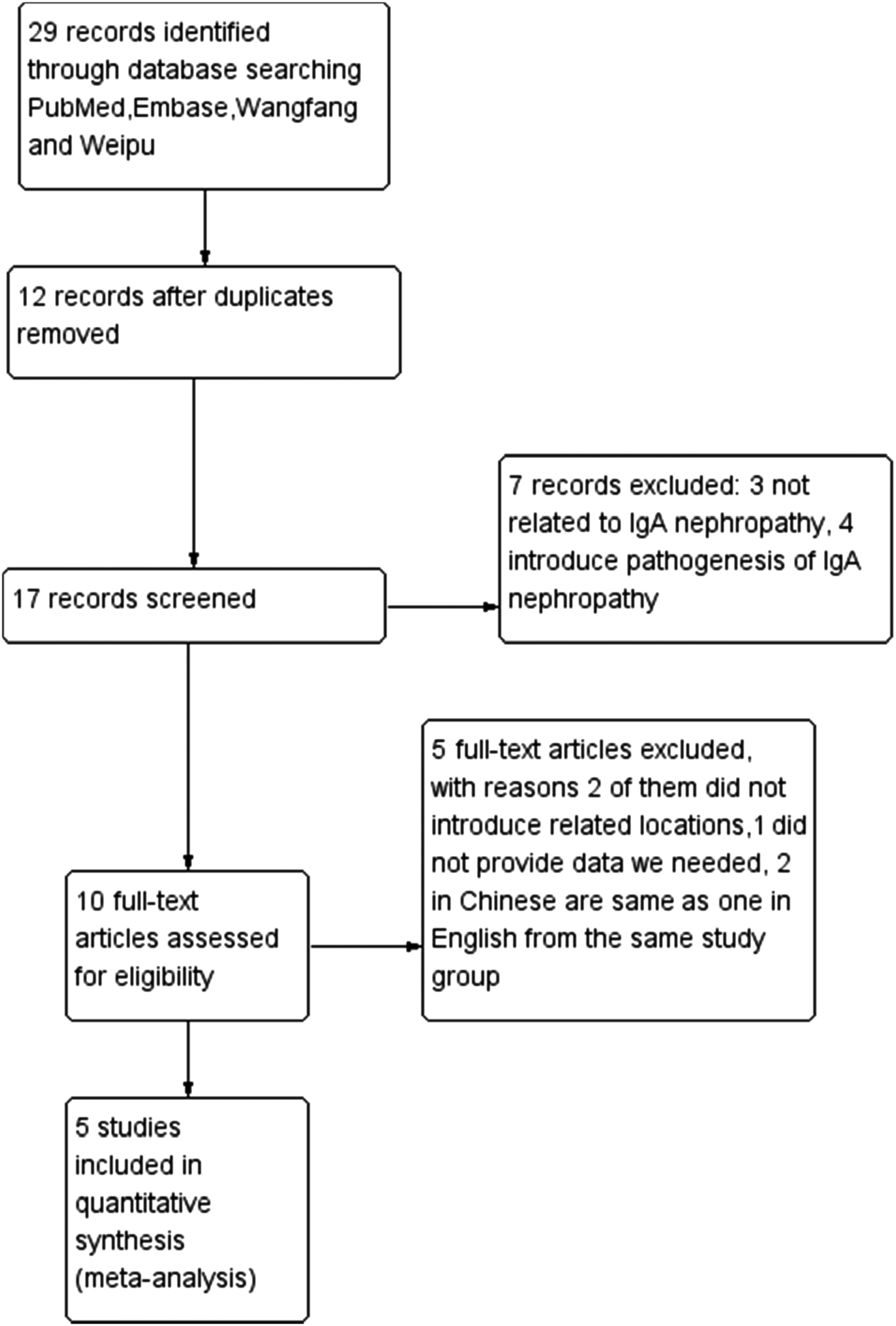
The process of selecting related studies.
Data extraction and analysis
The following information from each study was extracted independently by two investigators. First author's name, year of publication, country, ethnicity, sample size, genotype methods, source of the control groups, and the number of cases and control subjects for C1GALT1 genotypes. The information was compared and disagreements were resolved by discussion.
Statistical analysis
Stata/SE 12.0 (StataCorp LP, College Station, TX) was used to calculate and analyze the data from each study. The odds ratio (OR) and 95% confidence interval (CI) were used to evaluate the association of C1GALT1 SNPs rs1008898 and rs1047763 with IgAN risk. The heterogeneity of each included study was calculated by p value of Q test. The Rodman-effects model was used to pool ORs with dichotomous data, but if p > 0.1, then a fixed-effects model was utilized. A chi-square test was applied to calculate if the distributions of genotype in controls were consistent with the Hardy–Weinberg equilibrium (HWE). We considered a p value <0.05 as significant. If the control group did not conform to HWE, we performed a sensitivity test. In our meta-analysis, the p value of HWE in controls for rs1047763 was 0, which did not conform to HWE. The bias of publication was tested by Egger's test and Begg's test. For SNP rs1047763, the values of Egger's test and Begg's test were 0.069 and 0.308, respectively; for SNP rs1008898, the values of these tests were 0.679 and 1.000, respectively. Therefore, our enrolled studies did not display publishing bias (we used the standard of p > 0.05).
We adopted the standard of 50% to indicate that when the I2 < 50%, heterogeneity did not exist among studies. Evident heterogeneity was found in this meta-analysis.
Sensitivity analysis
We used Stata/SE 12.0 to perform sensitivity analysis and excluded each study every time.
Results
We used Stata/SE12.0 to analyze the associations of rs1047763 and rs1008898 with IgAN risk.
Study characteristics
We searched 29 studies from electronic databases: 11 from PubMed, 12 from EMBASE, 4 from Weipu, and 2 from Wanfang. We excluded repeated studies and studies with no related information regarding the pathogenesis of IgAN. If there were multiple publications of similar data by the same group, we enrolled the study with the largest sample size. If the same report appeared in English and Chinese, we enrolled the English language study.
There were five studies included in our meta-analysis (Pirulli et al.,(17) Li and Lu,(19) Mostowska et al.,(18) Li et al.,(20) and Wang et al.(6)), with four studies about rs1047763 (Li et al.(20) and Li and Lu(19) from China, Pirulli et al.(17) from Italy, and Mostowska et al.(18) from Poland) and three studies about rs1008898 (Li et al.,(20) Li and Lu,(19) and Wang et al.,(6) all of them from China). All five articles provided the related allele and genotype distributions we needed. The controls in these studies were tested for conformation to HWE; Pirulli et al.(17) (p = 0.049) and Li and Lu(19) (p = 0.045 and 0.014 for rs1047763 and rs1008898, respectively) were not in HWE. The remaining control groups in the enrolled studies were all in HWE. In the current meta-analysis, the control groups for rs1047763 (p = 0) and rs1008898 (p = 0.38) were not in HWE.
Allele and genotype distribution in the IgAN and healthy control group
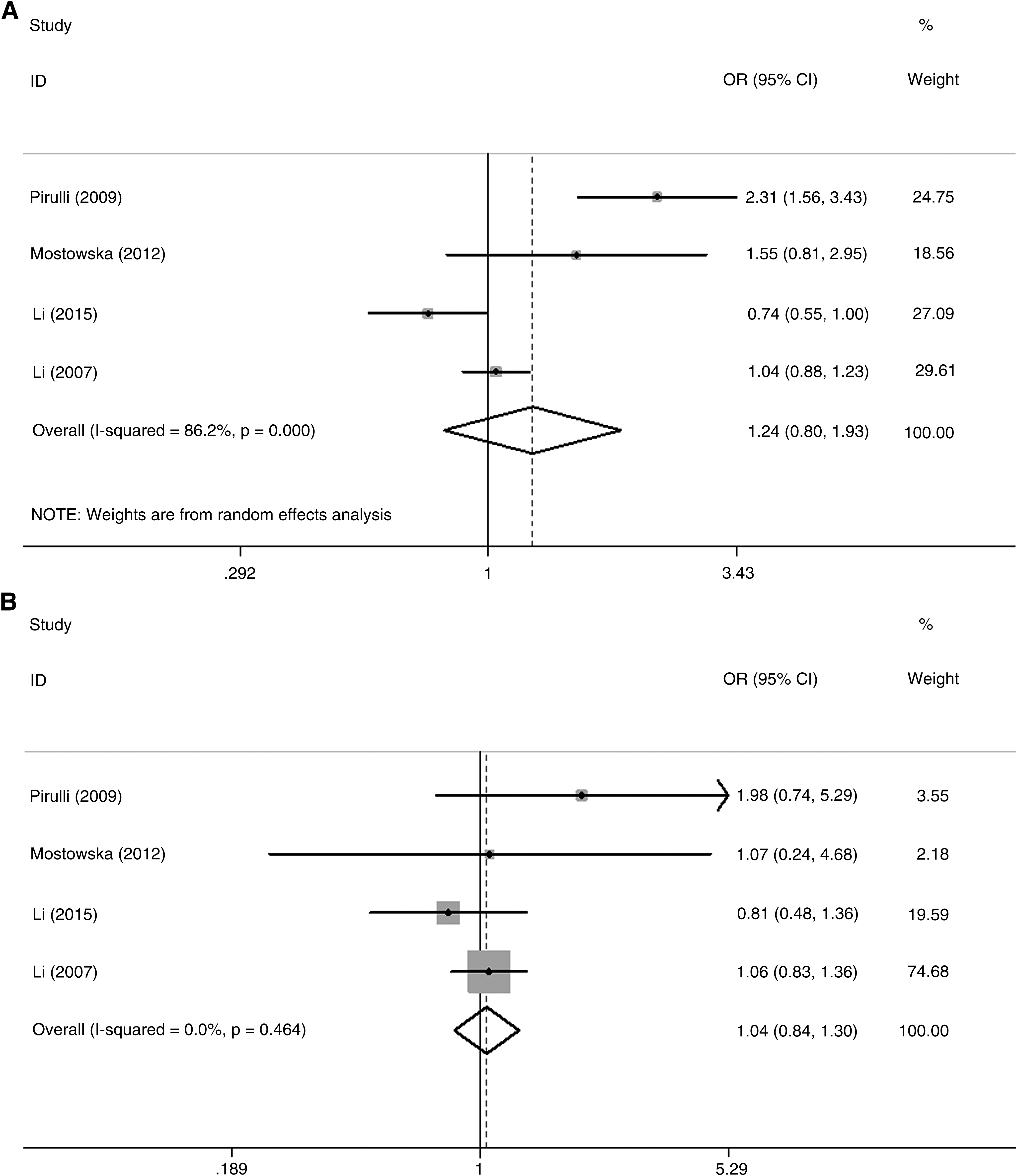
Our meta-analysis indicated that the C1GALT1 rs1047763 G allele and GG genotype were not associated with the risk of IgAN. GG, AG, and AA were found in 457 (39.21%), 452 (38.80%), and 256 (21.99%) IgAN patients, and in 556 (42.27%), 530 (40.22%), and 230 (17.51%) control subjects, respectively (G vs. A: OR = 1.244, 95% CI: 0.802–1.932, I2 = 86.2%, p = 0.330, Fig. 2A; GG+AG vs. AA: OR = 1.044, 95% CI: 0.840–1.297, I2 = 0, p = 0.699, Fig. 2B; GG vs. GA+AA: OR = 1.291, 95% CI: 0.702–2.374, I2 = 86.5%, p = 0.411, Fig. 2C). At the same time, we also performed a comparison between GG and AA genotypes, but no significant difference was found (OR = 1.019, 95% CI: 0.777–1.337, I2 = 47.9%, p = 0.890, Fig. 2D).
Our results also did not show any association between rs1008898 and the risk of IgAN. The number of GG, GT, and TT genotypes was 359 (26.07%), 643 (46.70%), and 375 (27.23%) in IgAN patients, and 341 (28.02%), 592 (48.64%), and 284 (23.34% in control subjects, respectively (T vs. G: OR = 1.159, 95% CI: 0.801–1.676, I2 = 89.5%, p = 0.434, Fig. 3A; TG+TT vs. GG: OR = 1.144, 95% CI: 0.693–1.888, I2 = 85.9%, p = 0.598, Fig. 3B; TT vs. GG+TG: OR = 1.241, 95% CI: 0.780–1.974, I2 = 81.0%, p = 0.361, Fig. 3C). Also, there were no statistically significance differences between TT genotype, GG genotype, and IgAN risk (OR = 1.271, 95% CI: 0.616–2.621, I2 = 88.6%, p = 0.516, Fig. 3D) (Tables 1–3).

HWE, Hardy–Weinberg equilibrium.
SNP, single-nucleotide polymorphisms.
Analysis of heterogeneity
When we conducted this meta-analysis, we found obvious heterogeneity in rs1047763 and rs1008898 (p value of I2 > 50%). We did not perform subgroup analysis due to the limitation in the number of enrolled articles. We considered ethnicity an important source of heterogeneity because there were four different ethnicities in rs1047763and three different ethnicities in rs1008898. In future studies, we expect greater sample numbers and multiple-center studies will overcome this problem.
Discussion
IgAN is the most prevalent glomerulonephritis throughout the world and the most common cause of ESRD in China. The clinical features of IgAN include microscopic hematuria, gross hematuria, and proteinuria. In the sera of IgAN patients, the level of Gd-IgA1 is higher than in healthy controls and other nephropathy groups,(21) which may indicate that Gd-IgA1 plays a key role in the progression of IgAN. IgAN is characterized by elevated serum IgA levels and deposition of IgA in mesangial cells. Complement component 3, IgG, or IgM is present in most biopsy samples,(22) which suggests that immunity reactions participate in the progression of the disease.
O-linked glycan locations are present in the hinge area of IgA1 molecules. It was reported that IgA1 was significantly associated with IgAN, and the location of O-linked glycans may play an important role in the native three-dimensional conformation required for normal physiological function.(23) Gd-IgA1 is a type of immune complex characterized by aberrant Gal-deficient glycosylation of O-linked glycans in the hinge area of IgA1. These aberrant glycosylation locations, when linked to GalNAc, may in turn lead to sialic deficiency in galactokinase. Gd-IgA1 may provoke an autoimmune response to immune complexes deposited in mesangial areas, stimulate mesangial cells and the complement system, and finally result in injury to the kidney.(23) Although O-linked glycans in the hinge region of IgA1 in healthy individuals are composed of GalNAc with β1,3-linked Gal, a majority of the IgA1 in the glomerular deposits in patients with IgAN is Gal deficient. Aberrantly glycosylated IgA1 was also detected in serum and in circulating immune complexes in IgAN patients, while it was rarely found in healthy controls.(24–27) The process of galactosylation of IgA1 relies on the molecular chaperone Cosmc and C1GALT1, so that downregulation of Cosmc and decreased activity of C1GALT1 can also lead to aberrant galactosylation of IgA1.
The C1GALT1 gene plays an important role in the pathogenesis of IgAN by regulating the production of the enzyme C1GALT1. The C1GALT1 gene may therefore be a candidate gene involved in the pathology of IgAN. C1GALT1 has been frequently reported to be associated with IgAN risk,(28–31) but conclusions have been inconsistent. Our meta-analysis aimed at a more reliable conclusion. We analyzed five reports focused on SNPs of C1GALT1 covering 1693 IgAN patients and 1864 healthy controls, and all the data needed were in the original publications. Our conclusion was that neither rs1047763 nor rs1008898 was associated with IgAN risk. This result was consistent with previous studies, with the exception of Pirulli et al.,(17) in which the G allele and GG genotype were indicated as risk factors for IgAN. In the Pirulli et al.(17) study, the control group was not in HWE. As there was no previous meta-analysis examining the association between C1GALT1 and IgAN, our results made sense to some extent.
However, our conclusion should be accepted cautiously for the following reasons. First, the number of studies in our meta-analysis was limited, the sample sizes were small,(18) and consequently, statistical power was reduced. Second, when we performed analysis of the relationship between rs1047763 and IgAN risk, heterogeneity was clearly evident in some genetic models. Given the limitation in the number of enrolled studies, we did not conduct subgroup analysis. Third, the control population for rs1047763 IgAN patients was not in HWE. Furthermore, our meta-analysis did not show any gene–environment interaction.
In conclusion, our meta-analysis did not find any significant associations of rs1047763 and rs1008898 with IgAN risk. However, the number of studies enrolled in the meta-analysis was small and more studies should be conducted in the future.
Footnotes
Author Disclosure Statement
No competing financial interests exist.