
Abstract
Ghrelin is an orexigenic peptide hormone that primarily regulates growth hormone secretion, food intake, and energy homeostasis. It has been shown to also play a role in numerous higher brain functions, such as the regulation of inflammation and cell proliferation. Ghrelin is the endogenous ligand of the growth hormone secretagogue receptor (GHSR), a G-protein-coupled receptor highly expressed in brain and detectable in some peripheral tissues. The wide distribution of ghrelin receptor and the number of tissues and cell types known to respond to ghrelin suggest that a number of systems may be affected by treatment with this hormone or its analogues. In this study, we characterized a new GHSR specific monoclonal antibody recognizing specifically the ghrelin receptor. This could be a useful tool for immunoassays aimed at obtaining insights into the physiological and pathological significance of the GHSR/ghrelin system.
Introduction
N
To date, the physiological functions of GHSR have been extensively studied and expanded to include: the release of various hormones such as growth hormone (GH), adrenocorticotropic hormone, cortisol, and prolactin(3); the modulation of food intake and energy metabolism(4); the influence on glucose and lipid metabolism(3); the regulation of gastrointestinal motility and secretion(5) and pancreatic function(6); the regulation of cell proliferation and survival(7,8); the attenuation of pro-inflammatory cascades and regulation of immune function that play important roles in aging and gastrointestinal homeostasis(9); and cell protection in nervous and cardiovascular systems.(10–12)
The GHSR gene can originate as two isoforms of the receptor: a full length isoform composed of seven transmembrane domains named GHSR1a and a truncated isoform with only five transmembrane domains named GHSR1b.(13,14) Although the two isoforms have been described, only GHSR1a is able to bind ghrelin or its analogs.(13,15) GHSR1a is a typical heterotrimeric G protein-coupled receptor (GPCR) containing 366 amino acids. As a member of GPCRs, GHSR1a contains seven transmembrane α-helix hydrophobic domains connected by three intra- and extracellular domains, beginning with an extracellular N-terminal domain and ending with an intracellular C-terminal domain.(16)
GHSR1b gene is comprehensively expressed in various tissues.(17) It is therefore reasonable to assume that this receptor possesses some unidentified biological functions. Indeed, GHSR1b decreases the cell surface expression of GHSR1a and acts as a repressor of the constitutive activity of GHSR1a when overexpressed in HEK-293 cells.(18) This finding indicates that GHSR1b may act as an endogenous modulator for GHSR1a constitutive activity,(12) but further studies are needed to investigate the physiological role of GHSR1b.
Many findings provide evidence that ghrelin may function as a survival factor for neuronal cells and offer a new perspective on the potential role of this peptide in neuronal injury. Ghrelin, even at rather low doses (10−13 M), protects hypothalamic neuronal cells from cell death by inhibiting apoptosis.(19) Moreover, ghrelin stimulates the ERK1/2 and PI3K/Akt pathways(20,21,19,22) and can exert its neuroprotective effects through stimulation of the protein kinase A and C pathways.(19)
At least some of the neuroprotective effects of ghrelin appear to be mediated through activation of GHSR1a. This receptor is found at highest concentrations in the pituitary and hypothalamus(13) and acts primarily at these sites to stimulate GH release, induce a positive energy balance by stimulating food intake, and decrease adipose usage through GH-independent mechanisms.(23) There is also abundant GHSR1a expression in other neuronal populations,(24) including the dentate gyrus of the hippocampus, the CA2 and CA3 regions of the hippocampus, the substantia nigra, the ventral tegmental area, and various thalamic and brain stem nuclei, including the dorsal raphe nucleus.(25) GHSR1a has recently been identified in the thoracic, lumbar, and sacral regions of the spinal cord(26) and expressed in sympathetic and autonomic preganglionic neurons. Our previous data demonstrated increasing levels of GHSR expression in primary developing neurons and a significant decrease in mature neurons at longer times in cultures. This transient behavior in both cortical and hippocampal neurons indicates a time-dependent manner modulation of this receptor that needs further investigation.(27)
The wide distribution of the ghrelin receptor and the number of tissues and cell types known to respond to ghrelin through a yet unidentified mechanism suggest that a number of systems may be affected with the treatment with this hormone or its analogue. Thus, it is imperative that we understand the diverse functions of ghrelin and its mechanisms of action, including the receptors involved, to develop specific and safe agonists or antagonists targeting the mechanism of interest.(28)
For this purpose, we produced a new monoclonal antibody (MAb) recognizing the ghrelin receptor which could be useful in immunoassays to obtain insights into the physiological and pathological significance of GHSR/ghrelin system. The MAb designed as 5C9H2 recognizes the C-terminal region of GHSR sequence. The added value of this MAb, as it can't bind the truncated isoform GHSR1b, is that it is able to distinguish between the two receptor isoforms.
Materials and Methods
GHSR C-terminal human sequence
The C-terminal amino acid sequence of human ghrelin receptor was analyzed by FASTA, from European Bioinformatics Institute, to exclude the existence of human proteins with significant sequence homology. A 23-mer sequence that lacks in the GHSR isoform 1b (here underscored) was selected.
MWNATPSEEPGFNLTLADLDWDASPGNDSLGDELLQLFPAPLLAGVTATCVALFVVGIAGNLLTMLVVSRFRELRTTTNLVRLWQYRPWNFGDLLCKLFQFVSESCTYATVLTITALSVERYFAICFPLRAKVVVTKGRVKLVIFVIWAVAFCSAGPIFVLVGVEHENGTDPWDTNECRPTEFAVRSGLLTVMVWVSSIFFFLPVFCLTVLYSLIGRKLWRRRRGDAVVGASLRDQNHKQTVKMLAVVVFAFILCWLPFHVGRYLFSKSFEPGSLEIAQISQYCNLVSFVLFYLSAAINPILYNIMSKKYRVAVFRLLGFEPF
Immunization protocol
Three male CD2F1 mice (Charles River), 7–12 weeks old, were immunized with peptide GHSR C-terminal (50 μg/mouse) emulsified in the same volume (100 μL/mouse) of incomplete Freund's Adjuvant (Sigma-Aldrich). Peptide was injected subcutaneously, followed by three booster injections at 2-week intervals. For priming, the emulsion was prepared with Complete Freund's Adjuvant. Mice were housed in appropriate animal care facilities and handled according to international guidelines for experiments with animals.
Generation of GHSR hybridomas
After three booster injections, test bleeds were assayed for positive reactions to peptide GHSR C-terminal by indirect enzyme-linked immunosorbent assay (ELISA). Spleen cells from immunized mice were fused to P3x63Ag8.653 mouse myeloma cells (Biological Bank, Istituto Nazionale per la Ricerca sul Cancro IST, Genova, Italy) in the presence of a 50% solution (wt/mL) of polyethylene glycol (mw 3350; Sigma-Aldrich) to produce monoclonal antibodies according to standard procedures.(29)
Cells were plated in 96-well plates (Corning-Costar) at 37°C in a humidified atmosphere in the presence of 5% CO2 and 95% air in RPMI 1640 medium supplemented with 10% FCS,
Enzyme-linked immunosorbent assay
ELISA 96-well plates (Immobilizer Streptavidin, Nunc Thermo Scientific) were coated with 1 μg/mL of peptide GHSR C-terminal biotinylated diluted in phosphate-buffered saline (PBS) at 4°C overnight. Plates were washed thrice with PBS +0.05% Tween-20 (Sigma-Aldrich) (PBST) and blocked with 5% bovine serum albumin (Sigma-Aldrich) in PBS at 37°C for 1 hour. Supernatants (50 μL) from hybridoma cultures, immune serum, or purified antibody diluted in PBS +0.5% bovine serum albumin were added and incubated at 37°C for 2 hours. Plates were washed thrice with PBST before incubation with secondary goat anti-mouse IgG antibody conjugated to alkaline phosphatase (Millipore, Bedford, MA) at 37°C for 1 hour. After another three washes with PBST, the plates were incubated with p-nitrophenyl phosphate (Sigma-Aldrich) diluted to 1 mg/mL in diethanolamine substrate buffer (Thermo Scientific, Waltham, MA) at 37°C for 30 minutes. The optical density at 405 nm was detected on a Victor3 plate reader (PerkinElmer).
Immunoprecipitation and western blot analysis
For immunoprecipitation experiments, cells were lysed with lysis buffer containing 20 mM Na2HPO4 (pH 7.2), 150 mM NaCl, 2 mM EGTA, 25 mM NaF, 1 mM Na3VO4, protease inhibition cocktail (Sigma-Aldrich), 1% Triton X-100, and 0.5% saponin (Sigma-Aldrich). Lysates were clarified by centrifugation at 15,900 g at 4°C for 30 minutes. The BCA Protein Assay Kit (Thermo Scientific) was used to determine protein concentrations.
Clarified lysates were incubated with MAb GHSR C-terminal (10 μg) or anti-GHSR commercial polyclonal antibody (LSBio; LS-C346069) (2 μg) at 4°C overnight, then bound to protein A/G agarose (Sigma-Aldrich) at 4°C for 2 hours. The agarose was washed thrice with lysis buffer, and proteins were released from the agarose by boiling in NuPAGE LDS Sample buffer (Invitrogen) for 5 minutes. The proteins were then subjected to sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to 0.45 mm nitrocellulose membranes (Invitrogen) at 30 V for 110 minutes at 4°C. After washing thrice with PBS +0.1% Tween and twice with distilled water, the blots were incubated at 4°C overnight with anti-GHSR purified commercial polyclonal (LSBio) or MAb specific for GHSR diluted in blocking solution (PBS +0.1% Tween +5% Skim milk). The membranes were washed thrice with PBS +0.1% Tween and incubated with goat anti-rabbit IgG (for polyclonal antibody) and with anti-mouse IgG (for MAb) conjugated to horseradish peroxidase (Bio-Rad) at room temperature (RT) for 45 minutes. Following three washes with PBS +0.1% Tween and two with distilled water, the antibody-reactive bands were visualized using chemiluminescence with ECL and exposure to film (Hyperfilm ECL; Amersham).
Quantitative immunofluorescence assay (In-Cell Western assay)
Cells were cultured in 96-well plates in complete medium culture and incubated at 37°C. The next day, the cells were fixed with 3.7% formaldehyde for 20 minutes, washed with PBS, and then permeabilized with Triton X-100 (0.1% in PBS) for 15 minutes. Cells were blocked with Odyssey blocking buffer (LI-COR Biosciences) for 90 minutes at RT. Cells were then incubated in a humid chamber overnight at 4°C with primary antibodies, anti-GHSR C-terminal diluted in Odyssey blocking buffer. After four washes with PBS +0.1% Tween-20, cells were incubated for 1 hour at RT with LI-COR IRDye 800 labeled secondary antibodies (1:800) and with CellTag 700 (1:500) for cell number normalization, diluted in Odyssey blocking buffer. CellTag 700 is a nonspecific cell stain that accumulates in both the nucleus and cytoplasm. Finally, cells were washed four times with PBS +0.1% Tween-20 before the scan of the plate by Odyssey® Infrared Imaging System (LI-COR Bioscience). Data were obtained from quadruplicate experiments.
Hippocampal neuron preparation
Hippocampal neurons were isolated from embryonic (E18) Sprague Dawley rats (Envigo, Bresso). Briefly, following animal sacrifice, brains were isolated, meninges removed from the medial aspect of the cerebral hemispheres, and then hippocampi dissected and kept at 4°C until process completion.
Tissue were then incubated with 2.5% trypsin for 15 minutes in a water bath at 37°C, then gently washed and kept in culturing media. Hippocampal dissociation was performed by repeatedly pipetting them up and down with a functionalized sterile Pasteur pipette. Following mechanical dissociation, cells were plated at desired density in neuronal plating medium, let to recover for 4 hours, and then put in compete neuronal culturing medium.
Immunocytochemical staining
Primary neurons at different stages of development (4, 9, and 18 days in vitro) were fixed in 4% paraformaldehyde and 4% sucrose at RT, for 10 minutes. Primary and secondary antibodies were applied in GDB buffer (30 mM phosphate buffer, pH 7.4, containing 0.2% gelatin, 0.5% Triton X-100, and 0.8 M NaCl) for 2 hours at RT. The confocal images were acquired with a Leica SPE confocal microscope, using a Nikon (Tokyo, Japan) 40 × objective with a sequential-acquisition setting at a resolution of 102461024 pixels. Each image was a z-series projection taken at 0.8 mm deep intervals. The following antibodies were used: polyclonal antibodies against VGlut-1 (1:1000 dilution) were from Synaptic System (Gottingen), and secondary antibody Alexa 546 was from Life Technologies.
siRNA analysis
To carry out GHSR selective silencing RNA experiments, we used commercially available probes for GHSR siRNA (Ambion selected predesigned siRNA; Life Technologies). Transfection was carried out using Lipofectamine (Life Technologies) following the manufacturer's protocol. Cells at the time of transfection were cultured on 16 mm diameter cover glass at 60% confluency. The following volumes of reagents were taken into consideration: total transfection volume 1 mL, siRNA 15 pMol, DNA e GFP 0.4 mg, and transfection agent 3 mL. Cells were exposed to transfection agent/siRNA/DNA complex for 1 hour. Following incubation, cells were washed in Dulbecco's medium without phenol red and, subsequently, cultured in standard conditions. Forty-eight hours later, cells were fixed in 4% paraformaldehyde and 4% sucrose at RT, for 10 minutes. Finally, immunocytochemical staining protocol was carried out, and cells were stained for GHSR.
Statistical analysis
Statistical significance between groups was determined by one-way analysis of variance; p < 0.05 was considered significant. All calculations were performed using GraphPad Prism 3.0 (GraphPad Software, La Jolla, CA).
Results
The purpose of this work was to produce and characterize a MAb specific for the C-terminal sequence of ghrelin human receptor, to develop a tool to assess its expression.
The human GHSR amino acid sequence was carefully analyzed and a 23-mer synthetic peptide that sequenced SQRKLSTLKDESSRAWTESSINT from the intracellular portion of the human GHSR protein was designed and selected as the immunogen. After immunization of CD2F1 mice, the titers of anti-GHSR antibodies in the mice sera were detected by ELISA. Mouse 3 with a higher anti-GHSR antibody titer was selected for hybridoma production (Fig. 1).
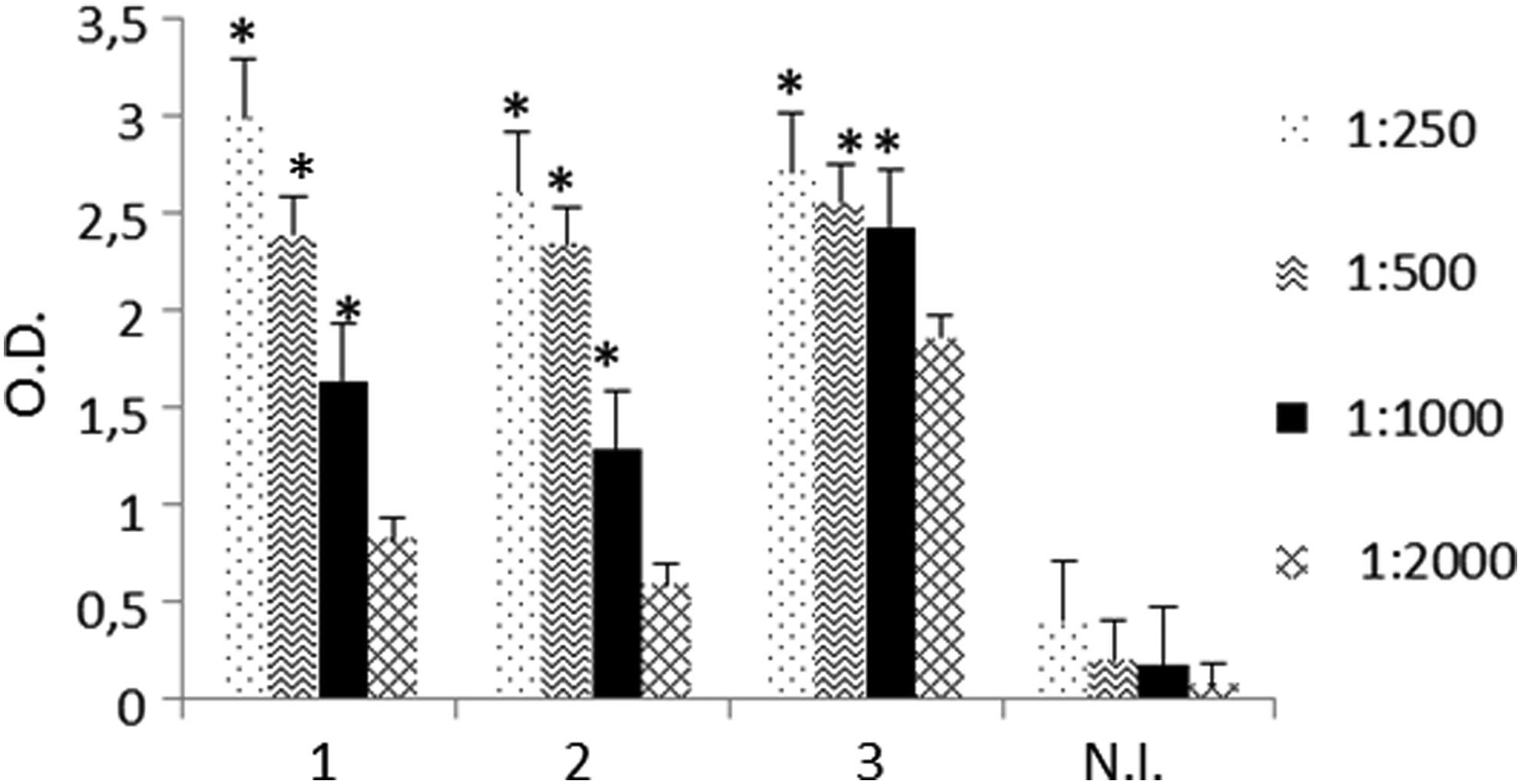
Antibody detection in sera from mice immunized with GHSR C-terminal peptide. The presence of specific antibodies for GHSR peptide in the sera of three immunized mice was determined by ELISA. Sera were used at different dilutions (1:250, 1:500, 1:1000, 1:2000). Serum from nonimmunized mice (N.I.) was used as a negative control. Results are expressed as mean ± standard error for three experiments. *p < 0.05 immunized vs. N.I. ELISA, enzyme-linked immunosorbent assay; GHSR, growth hormone secretagogue receptor.
After cell fusion with P3X63Ag8.653 mouse myeloma cells, supernatants of growing hybridoma cells were screened based on their reactivity with the peptide by ELISA. After cloning by limiting dilution, hybridomas showed stable murine monoclonal antibody anti-GHSR production. All clones were screened by ELISA and only anti-GHSR producing hybridomas whose supernatants responded with an optical density value 0.5 versus negative control were selected.
The monoclonal antibody subclass was determined as described in the Materials and Methods section. All the MAbs obtained were IgG (data not shown) and we selected the 5C9H2 hybridoma. After supernatant purification, we obtained a MAb with high specificity for its antigen (Fig. 2).
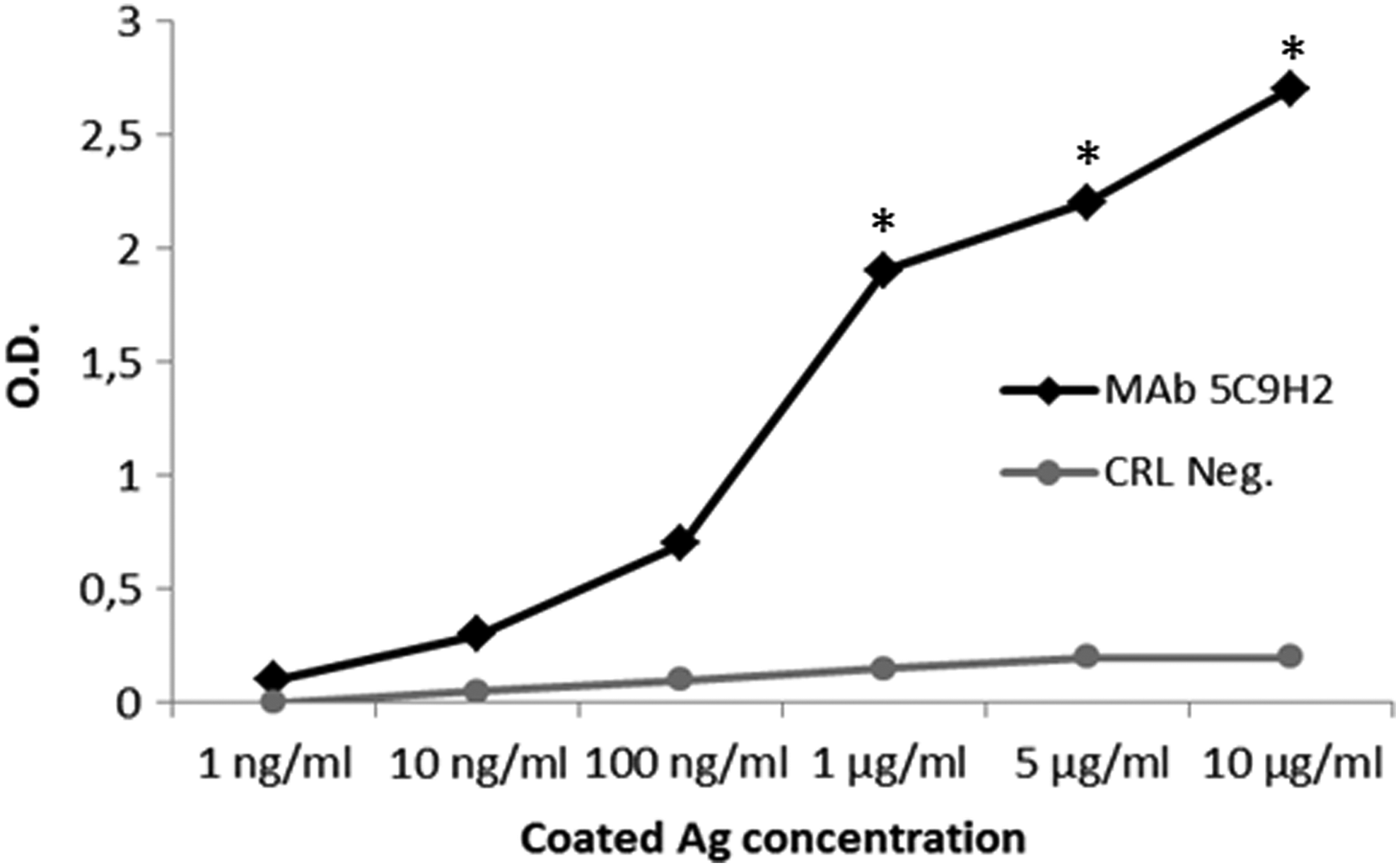
Specific binding analysis of MAb 5C9H2 for GHSR C-terminal peptide by ELISA. Peptide GHSR C-terminal was coated into microtiter plates, and serial dilutions of MAb 5C9H2 were added. The binding of MAb to antigens was detected with horseradish peroxidase-conjugated goat anti-mouse IgG. Results are expressed as mean ± standard error of three experiments. *p < 0.05 MAb 5C9H2 versus CRL Neg. MAb, monoclonal antibody.
First, to characterize the ability of the MAb to recognize GHSR, we immunoprecipitated protein extracts of 22RV1 cells, a human prostate cell line known to express the GHSR.(30) Lysates were immunoprecipitated with the selected MAb, and Western blot analysis was performed with a commercial anti-GHSR polyclonal antibody (Fig. 3a). In parallel, we immunoprecipitated 22RV1 cells with the commercial anti-GHSR polyclonal antibody and performed Western blot analysis with MAb 5C9H2 (Fig. 3b). In both cases, a 48 kDa protein band was detected, corresponding to the predicted size of GHSR, to indicate that our MAb is able to recognize the native GHSR protein in a specific manner.

Immunoprecipitation of 22RV1 cell lysates. Lysates from 22RV1 cells were immunoprecipitated with MAb 5C9H2
GHSR expression protein was also assessed in a quantitative immunofluorescence-based technique, an “In-Cell Western” assay (ICW). Results showed that MAb 5C9H2 was able to recognize GHSR in 22RV1 cells, and the decrease of signal on siRNA GHSR transfected cells confirmed the specificity of the binding (Fig. 4).
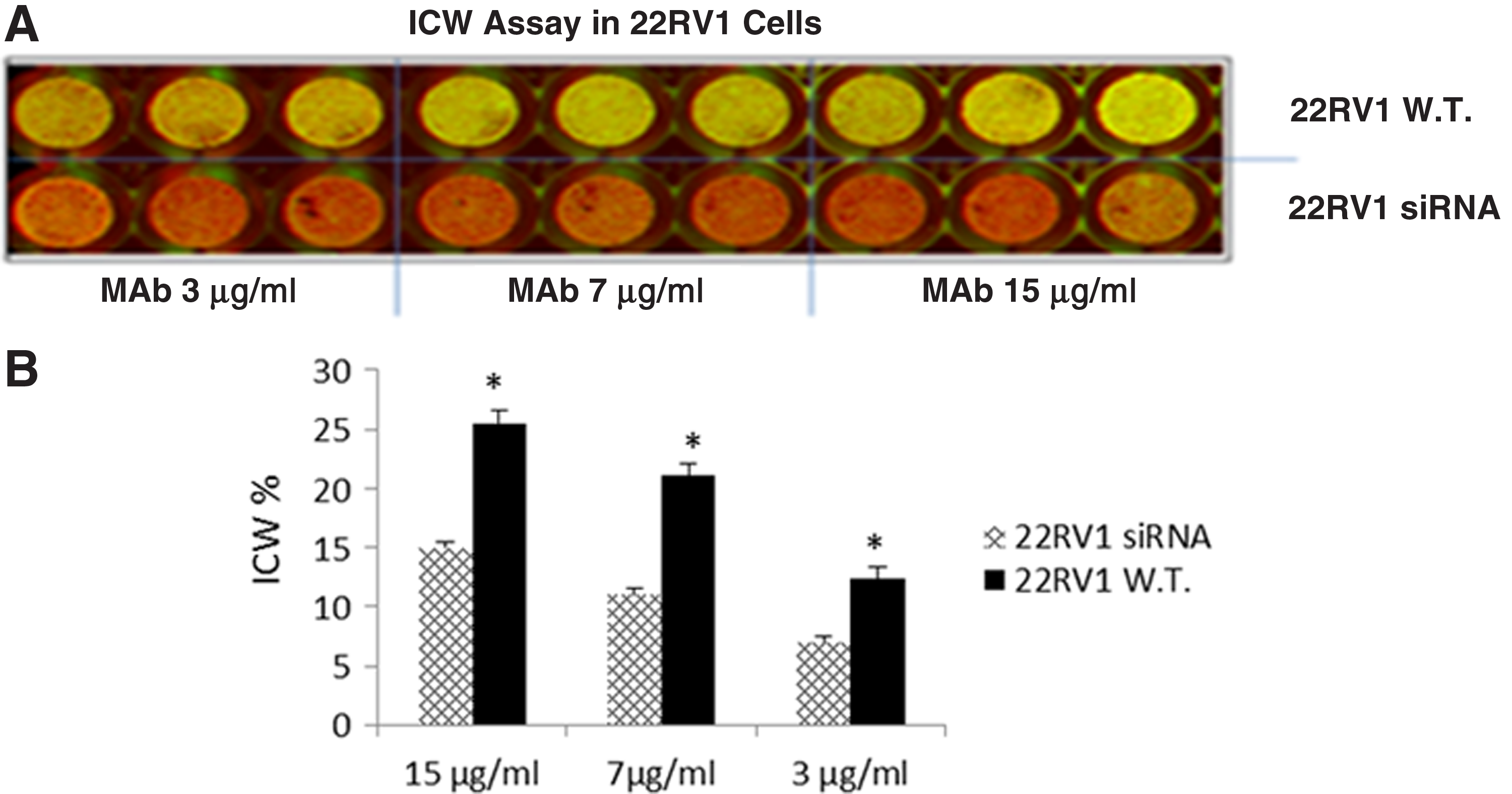
The detection of GHSR by 5C9H2 by In-Cell Western assay. 22RV1 wild type (22RV1 W.T.) and 22RV1 cells transfected with anti GHSR siRNA (22RV1 siRNA) were seeded in 96-well plate, fixed, and permeabilized. Cells were then incubated with MAb 5C9H2 at three concentrations, and the binding was detected with IRDye 800CW goat anti-mouse IgG antibody. CellTag 700 stain was also added in the wells to normalize the cell number.
At this point, we used our antibody to evaluate the GHSR expression in the central nervous system. More specifically, an immunoprecipitation of lysates of rat hippocampus was carried out with MAb 5C9H2, and Western blot analysis was performed with the commercial polyclonal antibody (Fig. 5).
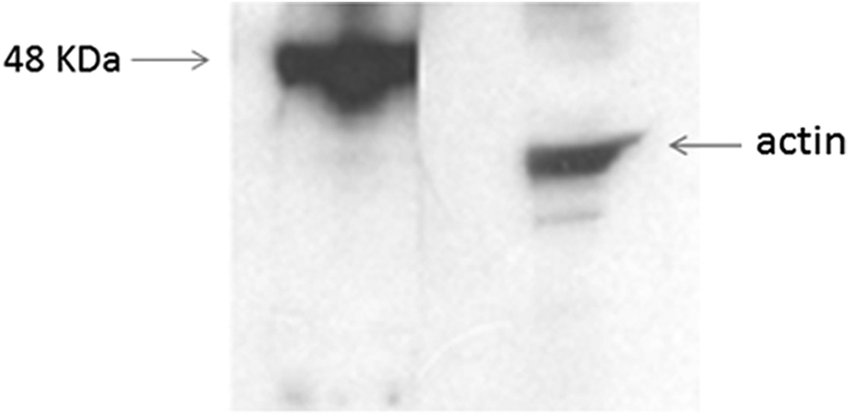
Immunoprecipitation of rat hippocampal primary neuron lysates with MAb 5C9H2. Lysates from rat hippocampal neurons after 9 days in vitro were immunoprecipitated with MAb 5C9H2, resolved, and transferred to nitrocellulose membranes. The western blot analysis was performed with the commercial anti-GHSR polyclonal antibody and clearly showed a 48 kDa band corresponding to the predicted size of GHSR. β-actin was used as a loading control.
We then proceeded by immunocytochemical staining of primary neurons at different stages of development, namely at 4, 10, and 18 days in vitro (div). These timings were selected because they represent the stages of ongoing neuronal development (4 div), maturation (10 div), and start of network activity and (18 div) mature neuronal network. Results indicated that the receptor is prevalently localized at the soma level in developing neurons and then it is distributed throughout the cell compartment, including dendrites and axon as the cell matures (Fig. 6).
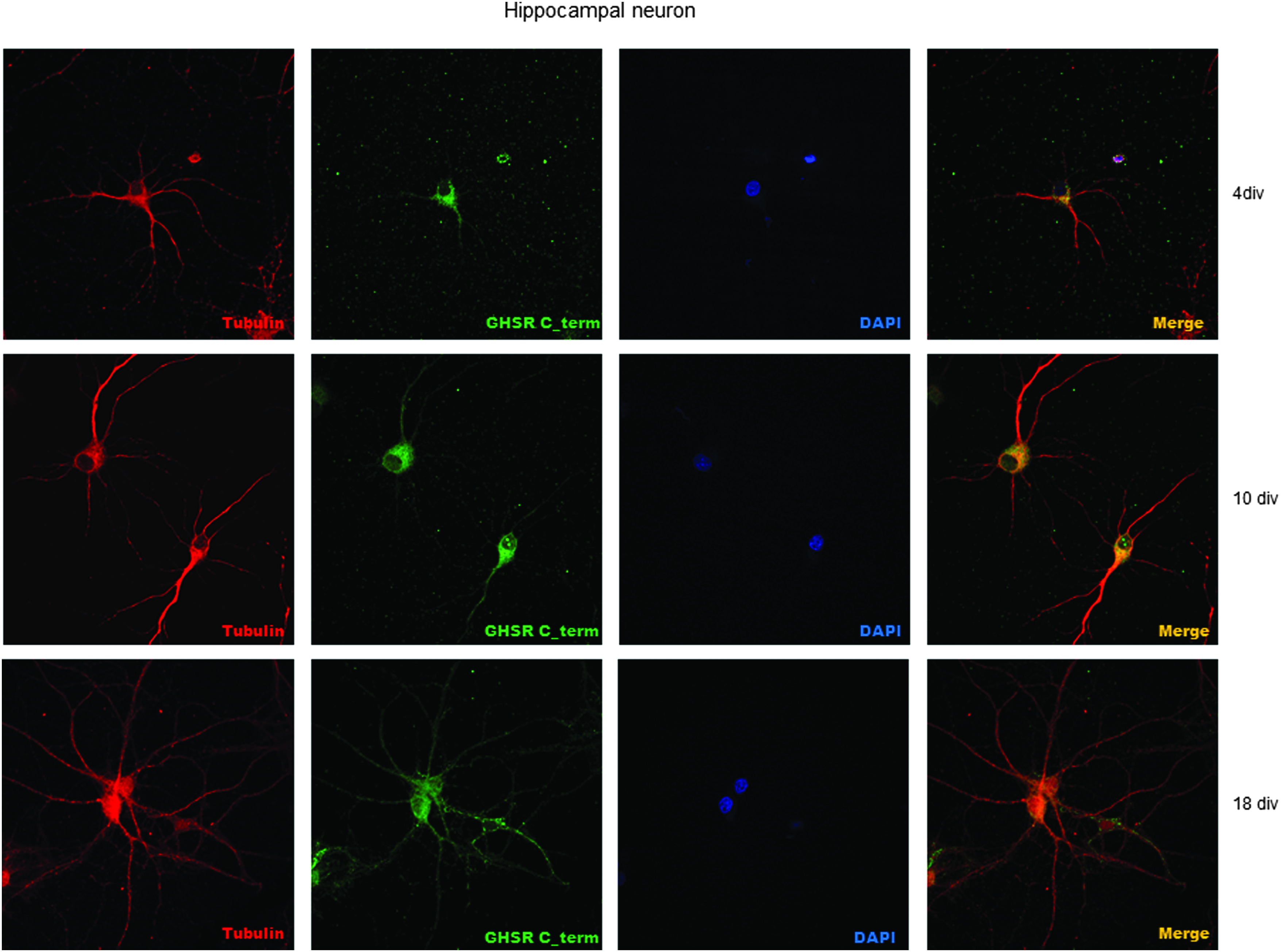
Immunocytochemical staining of primary rat hippocampal neurons at different stages of development (4, 10, 18 div). Cells were stained with cytoskeletal marker beta III tubulin, the newly developed MAb anti-GHSR and DAPI.
These results confirmed our previous observations and indicate not only that GHSR expression in primary neurons is modulated in a time-dependent manner but also that there might be a selective regional distribution which might yield to functional modulatory effects of ghrelin in the brain areas.(27)
Discussion
Increasingly numerous data underline the involvement of ghrelin and its receptors in several physiological and pathological processes. In this study we produced an anti-GHSR antibody, named 5C9H2, with IgG1 subclass to learn more about the GHSR/ghrelin system. Unlike all other commercial antibodies, 5C9H2 is a MAb specific for human and rat GHSR and our investigations confirm that it can be used in immunofluorescence, immunoprecipitation, and Western blot.
To immunize mice, we selected a 23-mer synthetic peptide from the intracellular portion of the human GHSR protein and FASTA analysis demonstrated that the chosen peptide did not share any homology with other human/rat protein. As this sequence is missing in GHSR1b, our MAb is able to distinguish between the two isoforms and binds only GHSR 1a, and ELISA revealed that the purified MAb 5C9H2 bound GHSR peptide in a specific concentration dependent manner (Fig. 2).
For the characterization of this antibody we used a line of human prostate cancer (22RV1), described as positive for GHSR.(30) Western blot analysis showed that MAb 5C9H2 was specific to GHSR human sequence and it was also able to recognize the sequence in its native state, as demonstrated by the immunoprecipitation of the GHSR protein from lysates derived from 22RV1 cells (Fig. 3a).
Moreover, the specificity of MAb 5C9H2 was demonstrated by silencing GHSR RNA in the prostate cell line and it was assessed with a quantitative immunofluorescence-based In-Cell Western assay which enables the detection of the antigen directly in the cells. We obtained a significant reduction of signal in siRNA GHSR transfected cells (Fig. 4).
Because there is abundant GHSR1a expression in different neuronal populations,(24) we then focused our attention on the brain tissue. We previous demonstrated(27) that there is a significantly higher expression of GHSR in hippocampal rather than cortical neurons, so an immunoprecipitation of rat hippocampus was carried out. In the Western blot analysis MAb 5C9H2 bound a 48 kDa protein band corresponding to the predicted size of GHSR, confirming its specific bind also in this tissue (Fig. 5).
The immunostaining of hippocampal neurons (Fig. 6) revealed that the receptor is present in the membrane of developed neurons and throughout its dendrites and axon, with a prevalent distribution in the first days of development at the somatic level.
In conclusion, the current study confirms the ability of our MAb 5C9H2 to recognize the GHSR protein. GHSR is involved in mediating a wide variety of biological effects of ghrelin and, dependent on the tissues and cells, the activation of this receptor may trigger a diversity of signaling mechanisms and subsequent distinct physiological responses. Therefore this MAb may provide a tool for future studies to better understand the cellular localization, as well as the functional role of this receptor.
Footnotes
Acknowledgments
The study was supported by Fernando Santarelli Foundation. Moreover, this work was partially supported by Consorzio Milano Ricerche.
Author Disclosure Statement
No competing financial interests exist.