
Abstract
Human lipocalin 6 (hLCN6) is a member of the lipocalin family, which is a group of structurally conserved hydrophobic ligand binding proteins, and widely distributed in animal, plant, and bacteria. Specific expression of hLCN6 in the epididymis and localization of this protein on the surface of spermatozoa suggest a role played by hLCN6, which may function as a transporter to carry ligands in the epididymal channel. However, the role of hLCN6 in sperm maturation has been largely unknown due to the lack of effective antibodies. In this study, we report the prokaryotic expression, purification, and refolding of recombinant hLCN6. Purified hLCN6 protein was used to generate monoclonal antibody (mAb) against this protein using conventional hybridoma techniques. The sensitivity and specificity of the anti-hLCN6 mAb were determined based on their activities in enzyme-linked immunosorbent assay and Western blotting analysis using various human tissues. The results showed that the antibody induced by recombinant hLCN6 protein had high sensitivity and specificity. Taken together, the recombinant hLCN6 protein and mAb against this protein obtained from our study provided useful tools for further exploration of the biological functions and molecular mechanism, as well as pathological significance of LCN6 in human.
Introduction
S
Human lipocalin 6 (hLCN6) is a newly discovered lipocalin family protein, sharing above 80% sequence homology with rat Lipocalin 6. The gene encodes a protein of 163 amino acids with a predicted signal peptide and a mature protein with a molecular weight of 16.0 kDa.(12) hLCN6 is a lipid carrier protein with a predicted basket-like β-barrel structure and high specificity of tissue distribution. It is predominantly expressed in the epididymis and located on the surface of spermatozoa, which is consistent with a role hLCN6 may play in spermatozoa maturation. The role could be to function as a transporter to carry ligands in the epididymal channel, a mechanism suggested for Lcn5 transport of retinoic acid.(13) When the expression of rat epididymal Lipocalin 6 was inhibited by RNA interference, the sperm motility decreased to less than 10% compared to the control (unpublished result). Therefore, Lcn6 plays a very important role for rat sperm maturation. Although the molecular mechanism of hLCN6 interaction with spermatozoa is unknown, its presence on spermatozoa in discrete dot-like distribution pattern raises the possibility of interaction with specific receptors.(14)
In the present study, to investigate the molecular mechanism of hLCN6 functions in vivo, we cloned and expressed the mature hLCN6 protein in Escherichia coli. The recombinant hLCN6 protein was then purified and re-natured and was used as antigen to generate mouse monoclonal antibody (mAb) by the hybridoma technology. Further characterization of the mAb showed that it could be used efficiently for detecting hLCN6 protein in human epididymal tissue. Our study provided useful tools for further exploration of the biological functions and pathological significance of hLCN6 in vivo.
Materials and Methods
Strains, culture media, chemicals, and antibodies
The plasmid vector pET28a and E. coli host strain DH5a and Rosetta (DE3) were purchased from CWBIO (Beijing, China). DNA Gel Extraction Kit and Plasmid Mini-prep Kit were from TIANGEN (Beijing, China). T4 DNA Ligase and restriction enzymes were products of Takara (Dalian, China). The tryptone and yeast extract were from Oxoid (Basingstoke, United Kingdom). DMEM and fetal bovine serum were products of GIBCO (Rockville, MD). Octaethylene glycol monododecyl ether (C12E8) was purchased from Sigma-Aldrich (Saint Louis, MO). The Ni-IDA and protein A agarose resin were products of GenScript (Piscataway, NJ). Other chemicals, except where indicated, were purchased from Sigma-Aldrich. Antibodies used in the study included anti-GAPDH mAb (Cell Signaling Technology) and horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG antibody (Boster Bio Tec, Wuhan, China). The mAb isotype control was obtained from SouthernBiotech (Birmingham, AL). BCA Protein Assay Kit and enhanced chemiluminescence (ECL) system and Amicon Ultra centrifugal filter were purchased from Pierce (Rockford, IL) and EMD Millipore (Darmstadt, Germany), respectively. Human tissues were obtained from the Forensic Expertise Center, Henan University of Science and Technology. All human tissues were obtained after family members' informed consent, and an institutional human research committee approved the investigation.
Construction of recombinant plasmid
Polymerase chain reaction (PCR) primers were designed to amplify the fragment encoding mature protein of hLCN6 (429 bp). The sequences of the primers were as follows: forward, 5′-ggaattc
Expression of recombinant protein in E. coli
E. coli Rosetta (DE3) was transformed with pET28a-hLCN6 recombinant plasmid. The transformed colony was cultured in 50 mL Luria-Bertani (LB) medium for 16 hours and then expanded in 3000 mL medium, and cells at the exponential stage were induced with 0.5 mM isopropyl-β-
Purification of the recombinant protein
Protein purification was performed with two methods. First is the method of denaturation and re-naturation.(15) Briefly, the Ni-IDA resin (5 mL) was added into a 15 mL tube and centrifuged at 800 g for 1 minute. The supernatant was gently removed, and the resin was washed with 10 mL water once and balanced with 10 mL denaturing binding buffer (50 mM Tris-Cl pH 8.0, 150 mM NaCl, 8 M urea). The pellet of the centrifuged cell lysate for 4 g of the cells was dissolved in 20 mL denaturing binding buffer. After centrifugation, 25 mL of dissolved pellet was mixed with the prewashed Ni-IDA resin, followed by incubation at 4°C for 15 hours while rotating. The mixture was then centrifuged at 800 g for 1 minute, and the supernatant was carefully removed and collected for the sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) analysis. The precipitate was eluted with 10 mL denaturing elution buffer I (50 mM Tris-Cl pH 8.0, 150 mM NaCl, 8 M urea, 50 mM imidazole) thrice, followed by a further elution with the denaturing elution buffer II (50 mM Tris-Cl pH 8.0, 150 mM NaCl, 8 M urea, 100 mM imidazole) with the same volume. Finally, the protein was eluted with 25 mL denaturing elution buffer III (50 mM Tris-Cl pH 8.0, 150 mM NaCl, 8 M urea, 500 mM imidazole). The fractions containing the purified target protein were combined and transferred into a dialysis bag and dialyzed successively against buffer IV (phosphate-buffered saline [PBS] containing 2 M urea, pH 7.4) at 4°C for 12 hours and buffer V (PBS, pH 7.4) for 5 hours before concentration.
The second method is detergent dissolution.(16) Briefly, the inclusion body was washed and then dissolved in 20 mL buffer A (50 mM Tris-Cl pH 8.0, 150 mM NaCl, 2% Triton X-100, 20 mM imidazole) at 4°C for 15 hours. After centrifugation, the supernatant was collected and mixed with 5 mL Ni-IDA resin prewashed by buffer A, followed by incubation at 4°C for 15 hours with rotating. The mixture was then centrifuged at 800 g for 1 minute, and the supernatant was carefully removed and collected for SDS-PAGE analysis. The precipitate was eluted with 10 mL washing buffer B (50 mM Tris-Cl pH 8.0, 150 mM NaCl, 2% Triton X-100, 500 mM imidazole) thrice. Fractions were manually collected at a flow rate of 1 mL/min. The eluted proteins were collected and analyzed by SDS-PAGE. The purified fractions were combined and concentrated to a final volume of ∼1 mL as described below before storing at −80°C.
The purified protein was centrifuged at 3000 g at 4°C for 30 minutes through the 10 kDa Amicon centrifugal filter to concentrate protein, and the concentration of the protein was determined using the BCA Assay Kit.(17) Proteins were analyzed using SDS-PAGE and Coomassie blue staining.
Preparation of mAb against human LCN6
Female BALB/c mice (7 weeks old) were immunized by subcutaneous (s.c.) injection with 100 μg of the purified hLCN6 obtained by the denaturation and refolding method in complete Freund's adjuvant. The subsequent three immunizations were carried out by s.c. injection at 2-week intervals with 100 μg of the same antigen mixed with the incomplete Freund's adjuvant. One week after the last immunization, the titer of anti-hLCN6 antibodies in the sera was monitored using enzyme-linked immunosorbent assay (ELISA). Three weeks later, a booster immunization was performed with an additional 100 μg of the same antigen in PBS by intraperitoneal (i.p.) injection. Three days after the last injection, splenocytes from the mice were collected and fused with SP2/0 myeloma cells in the logarithmic growth phase at the ratio of 20:1, followed by coculturing with peritoneal macrophages from normal BALB/c mice as feeder cells in 96-well plates.(18) Positive hybridomas were identified using indirect ELISA and cultured for a further 4 weeks, followed by subcloning by limiting dilution to obtain hybridoma cell lines stably secreting mAbs against human LCN6. The hybridoma clone with the highest ascite titer was expanded, and mAb was purified from the ascites using protein A agarose resin.
Identification of hLCN6 mAb isotype
The subtypes of the mAbs secreted by hybridoma cell lines were identified by sandwich ELISA using a mAb Isotyping Kit purchased from SouthernBiotech. The specific method was performed according to the kit instructions.
ELISA
Ninety-six well microplates were coated with 100 μL purified hLCN6 (5 μg/mL) in coating buffer (1.59 g Na2CO3 and 2.93 g NaHCO3 in 1 L H2O, pH 9.6) overnight at 4°C. After being washed twice with PBST (0.05% Tween-20 in PBS) and blocked with 3% bovine serum albumin (BSA) for 1 hour at 37°C, each well was incubated with 100 μL diluted antibody or culture supernatants in 3% BSA/PBST for an hour at 37°C. Negative control was incubated with serum from nonimmunized mice. One hundred microliters HRP-conjugated goat anti-mouse IgG antibodies in 3% BSA/PBST was added to each well for 1 hour followed by washing thrice with PBST. Plates were washed with PBST thrice, then 50 μL TMB solution was added to each well, and absorbencies were determined after 10 minutes at 450 nm by a microplate reader.
Western blotting
The tissues were fully triturated, extracted with radio-immunoprecipitation assay (RIPA) lysis buffer containing phenylmethanesulfonyl fluoride for 30 minutes on ice, and centrifuged. The protein concentration in the supernatants was determined using the bovine serum albumin (BSA) Protein Assay Kit. The protein samples were separated by SDS-PAGE and electrotransferred onto polyvinylidene fluoride membranes. After blocking at room temperature with 5% skim milk in PBST for 2 hours, the membranes were incubated with anti-hLCN6 mAb (1:10,000 dilution) at 4°C overnight. After washing, the membranes were incubated with secondary antibodies (1:2000 dilution) for 2 hours at room temperature and developed using the ECL reagents after extensive washing. The GAPDH was used as an internal control.
Results
Construction of recombinant plasmid to express hLCN6
To express the hLCN6 protein as antigen, we amplified the fragment encoding mature protein of hLCN6 (429 bp) using PCR. The PCR product was confirmed by agarose gel electrophoresis and was inserted into the pET28a vector. The resulting plasmid, pET28a-hLCN6, was further confirmed by PCR (Fig. 1) and DNA sequencing (data not shown). The construct was expected to express fusion protein containing an N terminal His tag and the mature hLCN6 protein.

Confirmation of recombinant plasmid expressing the mature hLCN6 protein. Two positive clones (lane 1 and lane 2) were screened and identified by PCR and then the amplified fragment encoding mature protein of hLCN6 was analyzed with agarose gel electrophoresis. The position of the amplified fragment is indicated by arrows. M, DNA molecular weight marker (from top, 15,000, 10,000, 7500, 5000, 2500, 1000, and 250 bp). hLCN6, human lipocalin 6; PCR, polymerase chain reaction.
Expression and purification of recombinant hLCN6 protein
E. coli Rosetta (DE3) was transformed with pET28a-hLCN6. One positive clone was selected and amplified in LB medium, and the expression of the recombinant protein was induced using IPTG in different temperatures. As shown in Figure 2A, a novel protein band with expected molecular weight was detected in E. coli transformed with pET28a-hLCN6. To purify the hLCN6 fusion protein, the IPTG-induced bacteria were harvested and resuspended. The cells were lysed with ultrasonic cytolysis and centrifuged to collect the supernatant and precipitate containing inclusion body, followed by the SDS-PAGE analysis with Coomassie blue staining. The result indicated that the majority of hLCN6 fusion protein derived from pET28a-hLCN6 was in the inclusion body fraction. We next purified the hLCN6 fusion protein in the inclusion body using affinity chromatography with the Ni2+-IDA column. A single protein band with the purity of 96% with the expected molecular weight was obtained, as shown by SDS-PAGE and Coomassie blue staining (Fig. 2B). Protein quantification indicated that 1.22 mg of hLCN6 fusion protein was obtained from 98 mg target protein in the inclusion body, with the overall percent recovery of 1.24%.
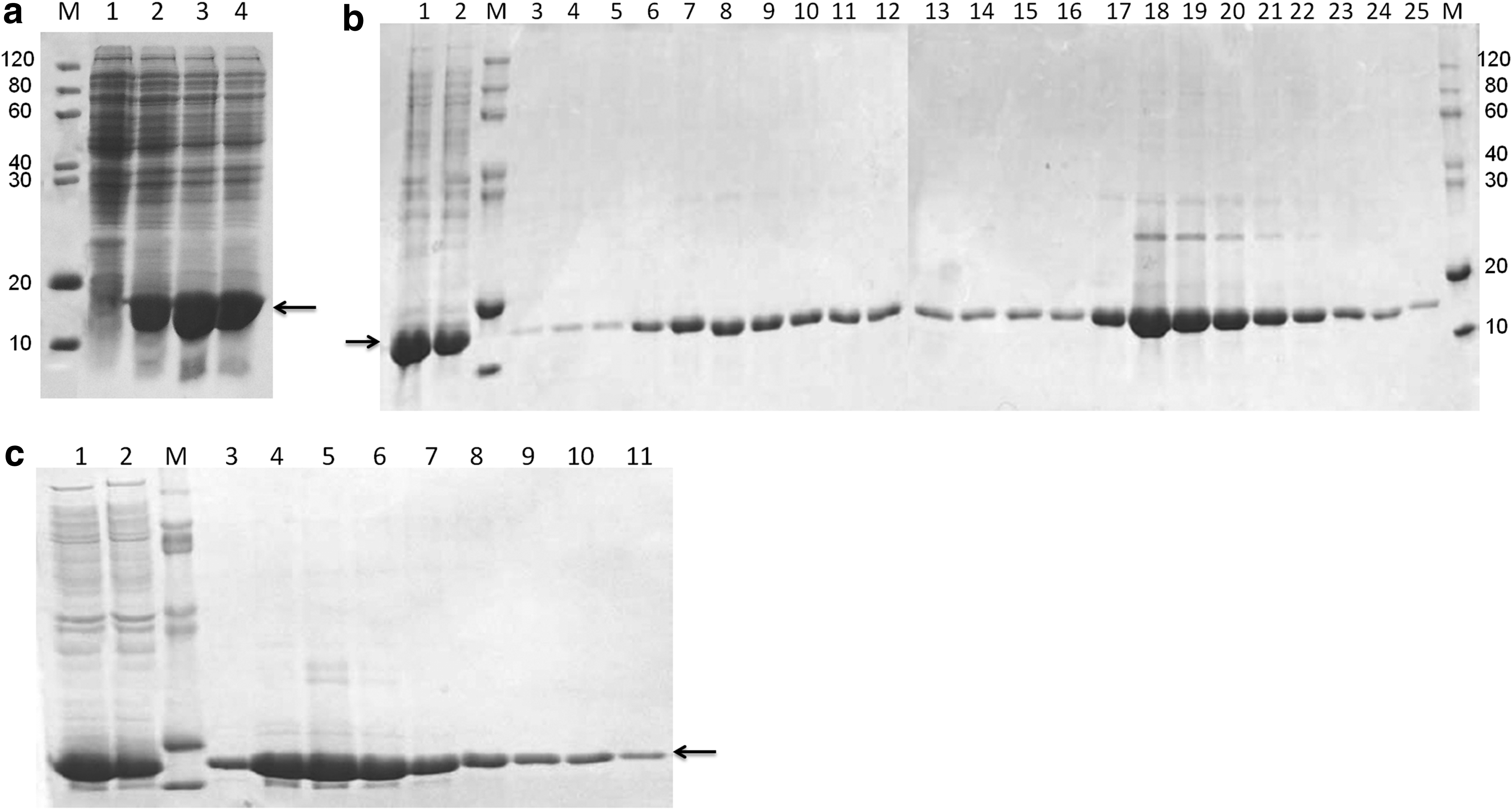
Expression and purification of the human LCN6 in Escherichia coli.
The hLCN6 fusion protein was also purified with the method of detergent dissolution. The target protein in the inclusion body could be partially dissolved in the buffer containing 2% Triton X-100. We further purified the hLCN6 fusion protein using affinity chromatography with the Ni-IDA column. A single protein band with the purity of 95% with the expected molecular weight was obtained, as shown by SDS-PAGE and Coomassie blue staining (Fig. 2C). Protein quantification indicated that 3.26 mg of hLCN6 fusion protein was obtained from 98 mg target protein in the inclusion body, with the overall percent recovery of 3.33%.
Identification of the recombinant hLCN6 protein
The fusion protein containing 6 × His tag was identified in SDS-PAGE (Fig. 3A) and Western blotting with anti-His antibody (Fig. 3B). The homogeneous protein was collected and concentrated to about 2 mg/mL in the absence of detergent due to its hydrophobic nature (Fig. 3C). From this figure, many amino acids could be identified in the hydrophobic zone. Several detergents were screened, and C12E8 was found to increase the protein solubility to about 3 mg/mL.
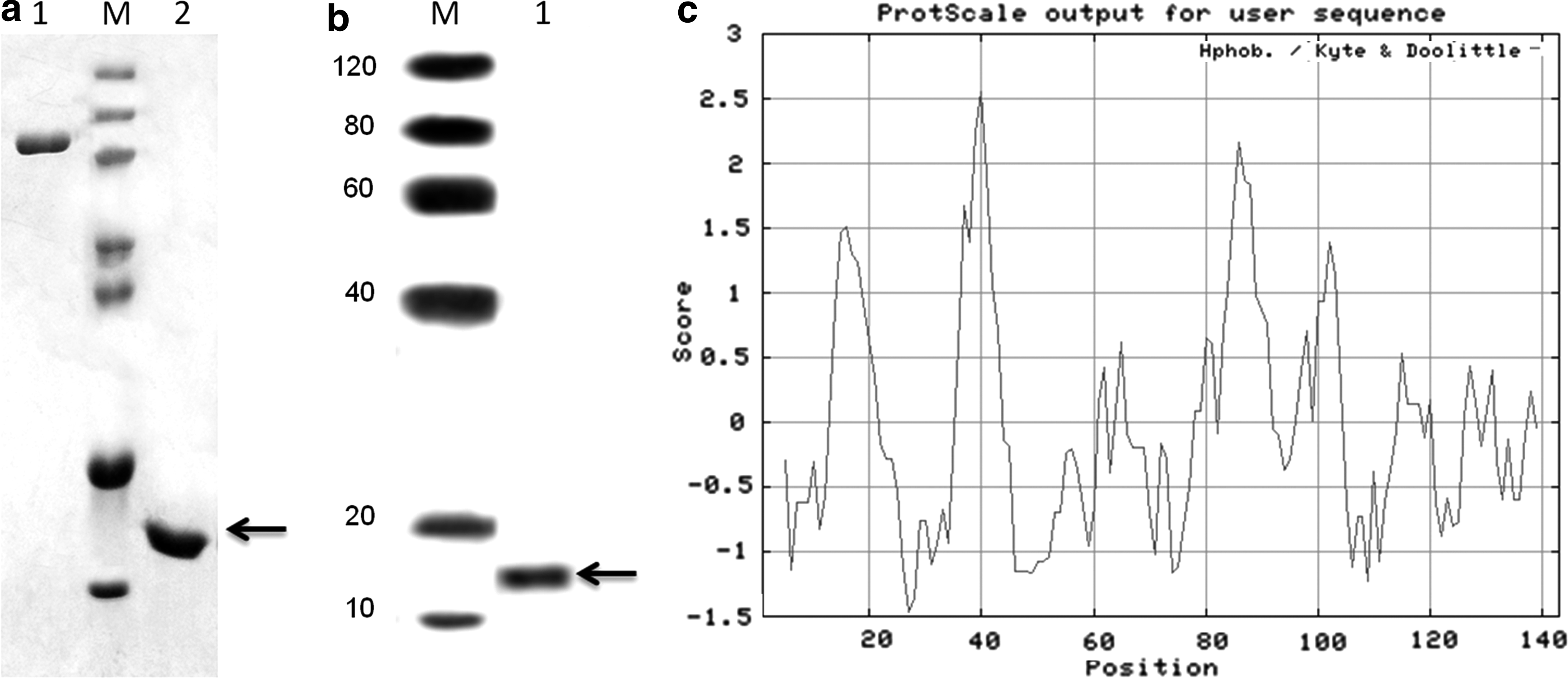
Identification of the recombinant hLCN6 protein. The purified and refolded hLCN6 fusion protein (∼16 kDa) was detected using SDS-PAGE
Generation of anti-mouse hLCN6 mAb
BALB/c mice were immunized with the purified hLCN6 protein according to standard procedures. Splenocytes were collected and fused with SP2/0 myeloma cells routinely, and positive hybridoma clones were screened using indirect ELISA. A total of eight hybridoma lines secreting anti-hLCN6 mAbs were established (Table 1). The isotypes of these mAbs were all characterized to be IgG1 + Kappa subclass. Ascites were prepared with one of these hybridomas named 1B2, and the titer of mAb purified from the ascites was determined by indirect ELISA. The titer of the purified anti-hLCN6 mAb was above 5.12 × 105 (Table 2). Then we identified the purified mAb by SDS-PAGE analysis with Coomassie blue staining. Two bands were detected corresponding to the heavy (55 kDa) and light (22 kDa) chains of the antibody (Fig. 4A). Next, we characterized the antigen-binding capability of the anti-hLCN6 mAb in Western blotting.
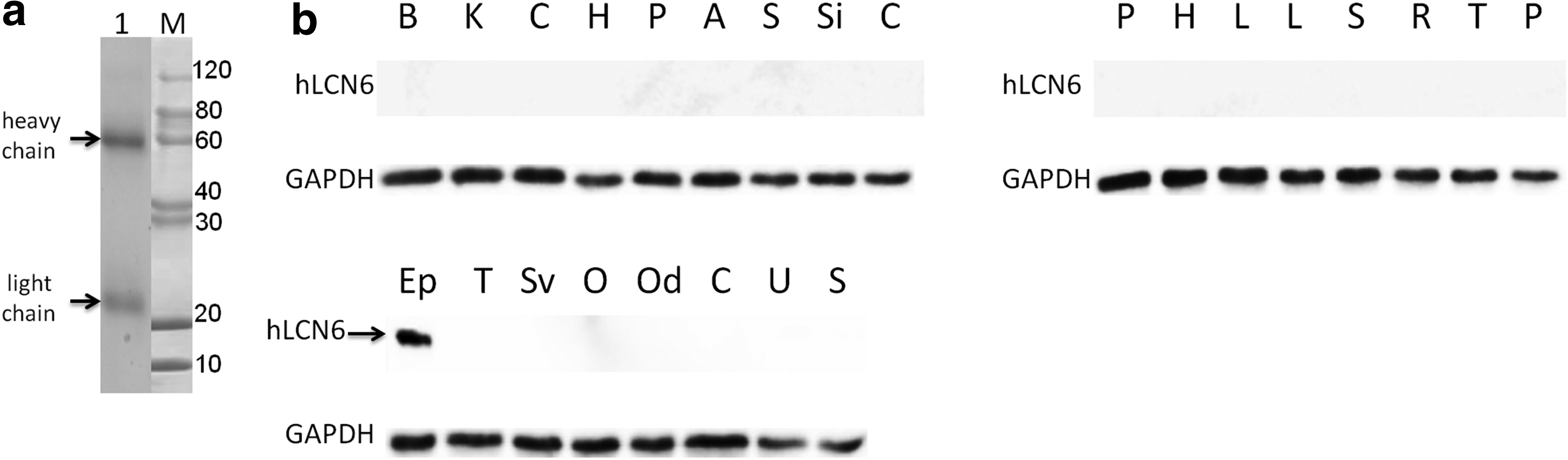
Characterization of the anti-human LCN6 mAb.
hLCN6 protein was exclusively expressed in epididymal tissue as identified by Western blotting using anti-hLCN6 mAb
To determine whether hLCN6 expression is predominantly in the epididymis as previously reported, protein extracts prepared from epididymis and 24 other human tissues were examined by Western blotting with the purified anti-hLCN6 mAb. The result showed that the hLCN6 antibody recognized a single band with a molecular weight of about 16 kDa only in a sample from the epididymis (Fig. 4B), suggesting that the hLCN6 antibody could bind to the mature hLCN6 protein in the Western blotting assay with high specificity. The result revealed that hLCN6 protein was exclusively expressed in human epididymal tissue. Moreover, the sensitivity of the anti-hLCN6 mAb was high since the signal could still be detected even if the primary antibody was diluted by 100,000 times (data not shown).
Discussion
A serious hurdle for structure determination in structure genomics is the acquisition of pure, soluble, and concentrated protein samples. Many proteins fail to adopt their three-dimensional structure and end up as inclusion bodies when expressed in the more conventional bacterial expression systems.(19) In this study, a successful protocol for hLCN6 expression, purification, and refolding was established. Anti-hLcn6 mAb was also obtained using conventional hybridoma technology.
For the expression of hLCN6, we found that almost all of the target proteins were expressed in insoluble inclusion body. Many attempts, involving decreasing the concentration of IPTG, expression at lower temperatures, and the use of various E. coli cell lines, were made to express the protein in the soluble fraction. The recombinant pET28a-hLCN6 protein was nevertheless expressed as inclusion body in cells. Therefore, we decided to purify the target protein by denaturation of the inclusion body and then refolding.
In view of the fact that some detergents can effectively enhance the solubility of the target protein, we tried to dissolve the inclusion body directly with detergent to obtain the hLCN6 protein. Nonionic detergents, which contain hydrophilic polar uncharged groups that do not disrupt protein–protein interactions, are frequently used for the separation and purification of membrane proteins. Therefore, we chose some commonly used nonionic detergents to dissolve the inclusion body. Experiments showed that Triton X-100 was effective in solubilizing hLCN6 protein partially from the inclusion body, and C12E8 was also able to dissolve a small amount of hLCN6 from the inclusion body (data not shown). To facilitate the subsequent purification of hLCN6 protein, we first dissolved the inclusion body partially with Triton X-100 and further purified the target protein from the supernatant with Ni-IDA affinity chromatography.
Although the hLCN6 protein was identified to express predominantly in the epididymis by Northern blot hybridization, the Western blotting analysis of hLCN6 expression has not been reported. In this study, using purified and refolded hLCN6 protein as antigen, we generated a set of mAbs against the human LCN6 protein. Antibody with considerably high avidity to the antigen was identified. We characterized the availabilities of these mAbs in ELISA and found that different clones of the mAbs exhibited different but close activities (Table 1). The anti-hLCN6 mAb with the highest activity was selected to make Western blotting analysis of hLCN6 expression in various human tissues. The result showed that hLCN6 protein was exclusively expressed in the epididymis, which was consistent with the Northern blot tissue distribution analysis.
On surfaces of cell types other than spermatozoa, several lipocalins interact with cell surface receptors.(20) The presence of hLCN6 on spermatozoa in a discrete dot-like distribution pattern raises the possibility of interaction with specific receptors. However, the molecular mechanism of hLCN6 interaction with spermatozoa is still unknown due to the lack of an effective mAb. In this study, we used purified and refolded recombinant hLCN6 protein to generate the anti-hLCN6 mAb with high sensitivity and specificity, providing a useful tool for identification of the potential sperm receptor interacting with hLCN6 and further for the exploration of the biological functions and molecular mechanism, as well as pathological significance of this protein in humans. The purified hLCN6 protein can also be used for crystallization and structure analysis. In addition, location of hLCN6 on sperm cell surface makes it a potential biomarker, and utilization of anti-hLCN6 mAb is reasonable for the separation of sperm cells from forensic mixture samples by immunobinding reaction.
Footnotes
Acknowledgments
The authors acknowledge Dr. Hua Diao for providing the recombinant pGEM-T Easy vector containing hLCN6 cDNA. This work was supported by grants from the National Natural Science Foundation of China (No. 81501634).
Author Disclosure Statement
No competing financial interests exist.