
Abstract
A type I transmembrane sialoglycoprotein podoplanin (PDPN) is expressed in several normal cells, including podocytes of the kidney, type I alveolar cells of the lung, and lymphatic endothelial cells. We recently produced an anti-bovine PDPN (bovPDPN) monoclonal antibody (mAb), PMab-44, by immunizing mice with recombinant proteins of bovPDPN. In this study, we determined the critical epitope of PMab-44 for the recognition of bovPDPN using many deletion mutants and point mutants of bovPDPN. Flow cytometric analyses revealed that the epitope of PMab-44 was Glu46-Thr50, which corresponds to platelet aggregation-stimulating (PLAG) domain-3. The important amino acids in the PMab-44 epitope were determined to be Glu46, Tyr48, and Thr50. Western blot analysis also confirmed these results, indicating that the PLAG domain of bovPDPN is also important in immunogenicity for producing useful anti-PDPN mAbs.
Introduction
A
PDPN is expressed in several normal cells, including lymphatic endothelial cells, renal podocytes, pulmonary type I alveolar cells, osteocytes, and chondrocytes.(10) The expression of PDPN has been reported in many malignant tumors, such as oral squamous cell carcinomas,(11) malignant brain tumors,(12–15) lung cancers,(16) esophageal cancers,(17) malignant mesotheliomas,(18,19) testicular tumors,(20) osteosarcomas,(10,21,22) and chondrosarcomas.(10) PDPN expression is also associated with malignant progression and cancer metastasis.(2,12,23)
We had previously established the anti-bovine PDPN (bovPDPN) monoclonal antibody (mAb) PMab-44 (IgG1, kappa), which was useful for detecting podocytes and lymphatic endothelial cells in normal bovine tissues in immunohistochemistry.(24) PMab-44 also detected bovPDPN specifically in flow cytometry. Our previous research revealed that bovPDPN lacks platelet aggregation-inducing activity because the predicted amino acid sequence of bovPDPN has a sporadic deletion in the PLAG domain.(25) Therefore, the functional character of bovPDPN may be somewhat different from other mammalian PDPN. However, bovPDPN has so far attracted little attention apart from a few studies.(26,27)
In this study, we further determined the epitope of PMab-44 in flow cytometry and western blotting. This information could be useful in bovPDPN veterinary research.
Materials and Methods
Cell lines
Chinese hamster ovary (CHO)-K1 was purchased from the American Type Culture Collection (ATCC, Manassas, VA). CHO-K1 cells were transfected with the bovPDPN-MAP tag(28) plasmid using a Gene Pulser Xcell electroporation system (Bio-Rad Laboratories, Inc., Berkeley, CA). CHO-K1 and CHO/bovPDPN were cultured in RPMI 1640 medium (Nacalai Tesque, Inc., Kyoto, Japan), supplemented with 10% heat-inactivated fetal bovine serum (Thermo Fisher Scientific, Inc., Waltham, MA), 100 units/mL of penicillin, 100 μg/mL of streptomycin, and 25 μg/mL of amphotericin B (Nacalai Tesque, Inc.) at 37°C in a humidified atmosphere of 5% CO2 and 95% air.
Production of bovPDPN mutants
The amplified bovPDPN cDNA was subcloned into a pCAG vector (Wako Pure Chemical Industries Ltd., Osaka, Japan) and a MAP tag(28) was added at the N-terminus. Deletion mutation of a bovPDPN sequence was performed using a HotStar HiFidelity polymerase chain reaction (PCR; Qiagen, Inc., Hilden, Germany) with oligonucleotides containing the desired mutations. PCR fragments bearing the desired mutations were inserted into a pCAG vector using the In-Fusion PCR Cloning Kit (Clontech, Palo Alto, CA). Substitutions of amino acids to alanine in bovPDPN sequences were conducted using the QuikChange lightning site-directed mutagenesis kit (Agilent Technologies, Inc., Santa Clara, CA). CHO-K1 cells were transfected with the plasmids using a Gene Pulser Xcell electroporation system (Bio-Rad Laboratories, Inc.).
Flow cytometry
Cells were harvested by brief exposure to 0.25% trypsin/1 mM EDTA (Nacalai Tesque, Inc.). After washing with 0.1% BSA/PBS, the cells were treated with PMab-44 (10 μg/mL) and anti-MAP tag(28) (clone: PMab-1, 1 μg/mL) for 30 minutes at 4°C followed by treatment with Oregon green-conjugated anti-mouse IgG or anti-rat IgG (1:1000 diluted; Thermo Fisher Scientific, Inc.). Fluorescence data were collected using a Cell Analyzer EC800 (Sony Corp., Tokyo, Japan). PMab-1 was previously developed against mouse PDPN(29) and has been utilized as an anti-MAP tag mAb.(28) PMab-1 does not crossreact with other PDPN proteins such as bovPDPN,(24) dog PDPN,(30–32) rat PDPN,(33) rabbit PDPN,(34,35) and human PDPN.(29)
Western blot analyses
Cell lysates (10 μg) were boiled in sodium dodecyl sulfate (SDS) sample buffer (Nacalai Tesque, Inc.). The proteins were electrophoresed on 5%–20% polyacrylamide gels (Wako Pure Chemical Industries Ltd.) and were transferred onto a PVDF membrane (Merck KGaA, Darmstadt, Germany). After blocking with 4% skim milk (Nacalai Tesque, Inc.), the membrane was incubated with PMab-44, anti-MAP tag(28) (clone: PMab-1), and anti-β-actin (clone AC-15; Sigma-Aldrich Corp., St. Louis, MO) and then with peroxidase-conjugated anti-mouse or anti-rat antibodies (1:1000 diluted; Dako; Agilent Technologies, Inc.). Subsequently, the membrane was developed with the Pierce Western Blotting Substrate Plus (Thermo Fisher Scientific, Inc.) or the ImmunoStar LD Chemiluminescence Reagent (Wako Pure Chemical Industries Ltd.) using a Sayaca-Imager (DRC Co. Ltd., Tokyo, Japan).
Determination of the binding affinity using flow cytometry
CHO/bovPDPN (2 × 105 cells) were resuspended with 100 μL of serially diluted antibodies (0.6 ng/mL–10 μg/mL) followed by secondary anti-mouse IgG (Thermo Fisher Scientific, Inc.). Fluorescence data were collected using a cell analyzer (EC800; Sony Corp.). The dissociation constants (KD) were obtained by fitting the binding isotherms using the built-in one-site binding models in GraphPad PRISM 6 (GraphPad Software, Inc., La Jolla, CA).
Results and Discussion
Epitope mapping of an anti-bovPDPN mAb PMab-44 by flow cytometry
We previously showed the usefulness of an anti-bovPDPN mAb PMab-44 in immunohistochemical analysis(24); in this study we performed the epitope mapping using flow cytometry and western blot analysis.
First, we produced five N-terminal deletion mutants of bovPDPN: dN23 (23–169 amino acids), dN40 (40–169 amino acids), dN60 (60–169 amino acids), dN80 (80–169 amino acids), and dN100 (100–169 amino acids). Flow cytometric analysis revealed that PMab-44 reacted with dN23 and dN40; in contrast, it did not react with dN60, dN80, and dN100 (Fig. 1A). The anti-MAP tag PMab-1 recognized all deletion mutants. These results indicate that the N-terminal of the PMab-44 epitope exists between Lys40 and Ala60 of bovPDPN.(25) Furthermore, we produced three N-terminal deletion mutants of bovPDPN: dN45 (45–169 amino acids), dN50 (50–169 amino acids), and dN55 (55–169 amino acids). Flow cytometric analysis revealed that PMab-44 reacted with dN23 and dN45; in contrast, it did not react with dN50 and dN55 (Fig. 1B). The anti-MAP tag PMab-1 recognized all deletion mutants. These results indicate that the N-terminal of the PMab-44 epitope exists between Val45 and Thr50 of bovPDPN.(25)
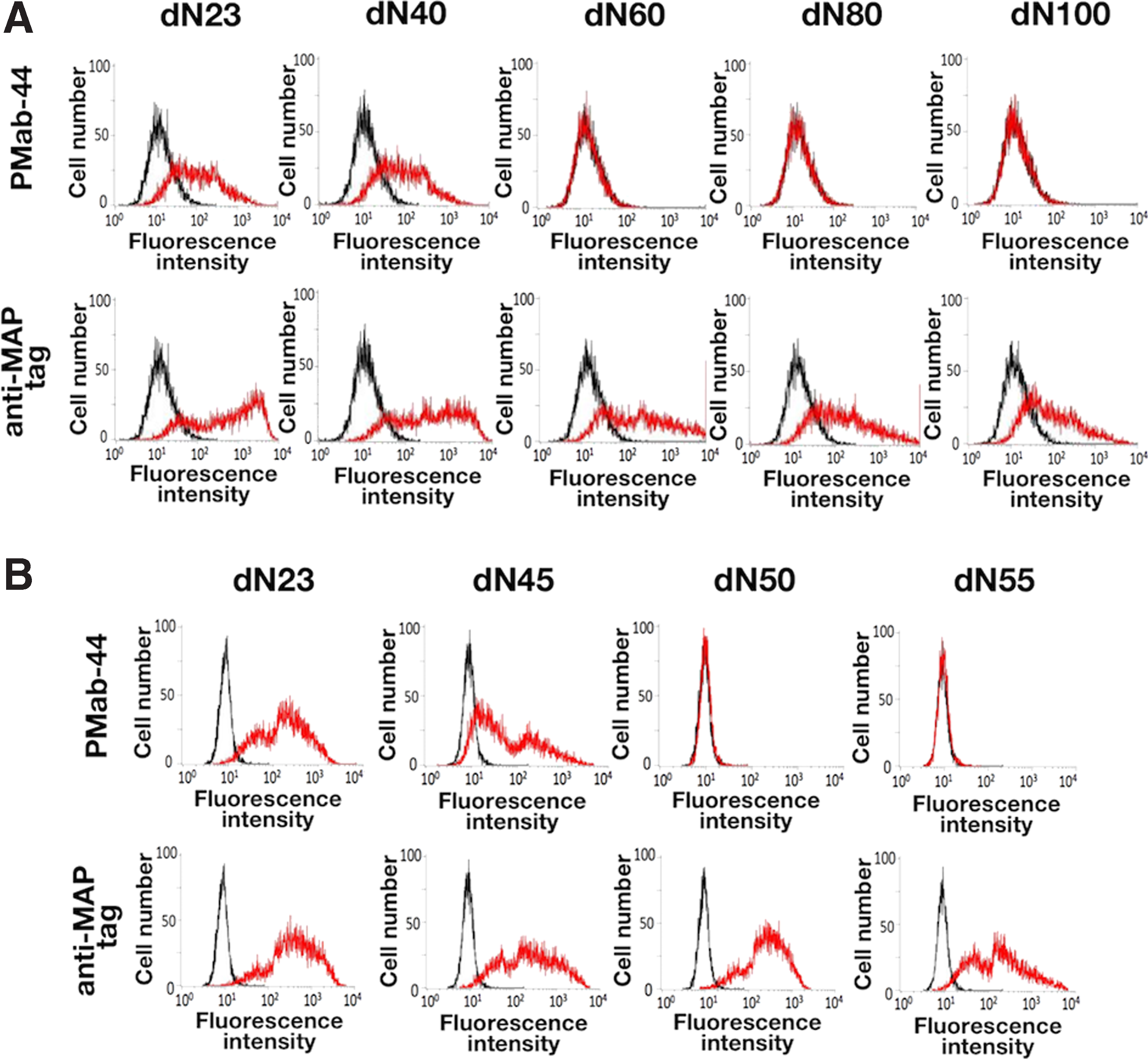
Epitope mapping of an anti-bovPDPN mAb PMab-44 using deletion mutants by flow cytometry.
Next, we produced 10 point mutants of bovPDPN: V45A, E46A, D47A, Y48A, T49A, T50A, T51A, P52A, A53G, and A54G. Figure 2 shows that two point mutants, such as E46A andY48A, completely lost the reaction by PMab-44, and T50A showed little reaction by PMab-44. All point mutants were recognized by the anti-MAP tag PMab-1. These results indicate that three amino acids of bovPDPN, Glu46, Tyr48, and Thr50, are critical for PMab-44 recognition.

Epitope mapping of an anti-bovPDPN mAb PMab-44 using point mutants by flow cytometry. Point mutants of bovPDPN (V45A, E46A, D47A, Y48A, T49A, T50A, T51A, P52A, A53G, and A54G) were treated with PMab-44 (10 μg/mL) and anti-MAP tag (1 μg/mL) for 30 minutes at 4°C followed by treatment with Oregon green-conjugated anti-mouse IgG or anti-rat IgG. Fluorescence data were collected using a Cell Analyzer EC800. Red line: PMab-44 or anti-PA tag. Black line: negative control.
Epitope mapping of an anti-bovPDPN mAb PMab-44 by western blot analysis
We next performed western blot analysis to confirm the results of epitope mapping of PMab-44 by flow cytometry. As shown in Figure 3A, PMab-44 did not react with V45A, E46A, D47A, Y48A, T49A, and T50A of bovPDPN. In contrast, PMab-44 reacted with T51A, P52A, A53G, and A54G. All point mutants were detected by the anti-MAP tag PMab-1. These results indicated that Val45, Glu46, Asp47, Tyr48, Thr49, and Thr50 are important in the recognition of PMab-44 against bovPDPN in western blot analysis. The epitope of PMab-44 is illustrated in Figure 3B. The sequence of the PMab-44 epitope (VEDYTT) shows low homology with the corresponding peptide sequence of human PDPN (AEDDVV), indicating that PMab-44 does not react with human PDPN.(24)
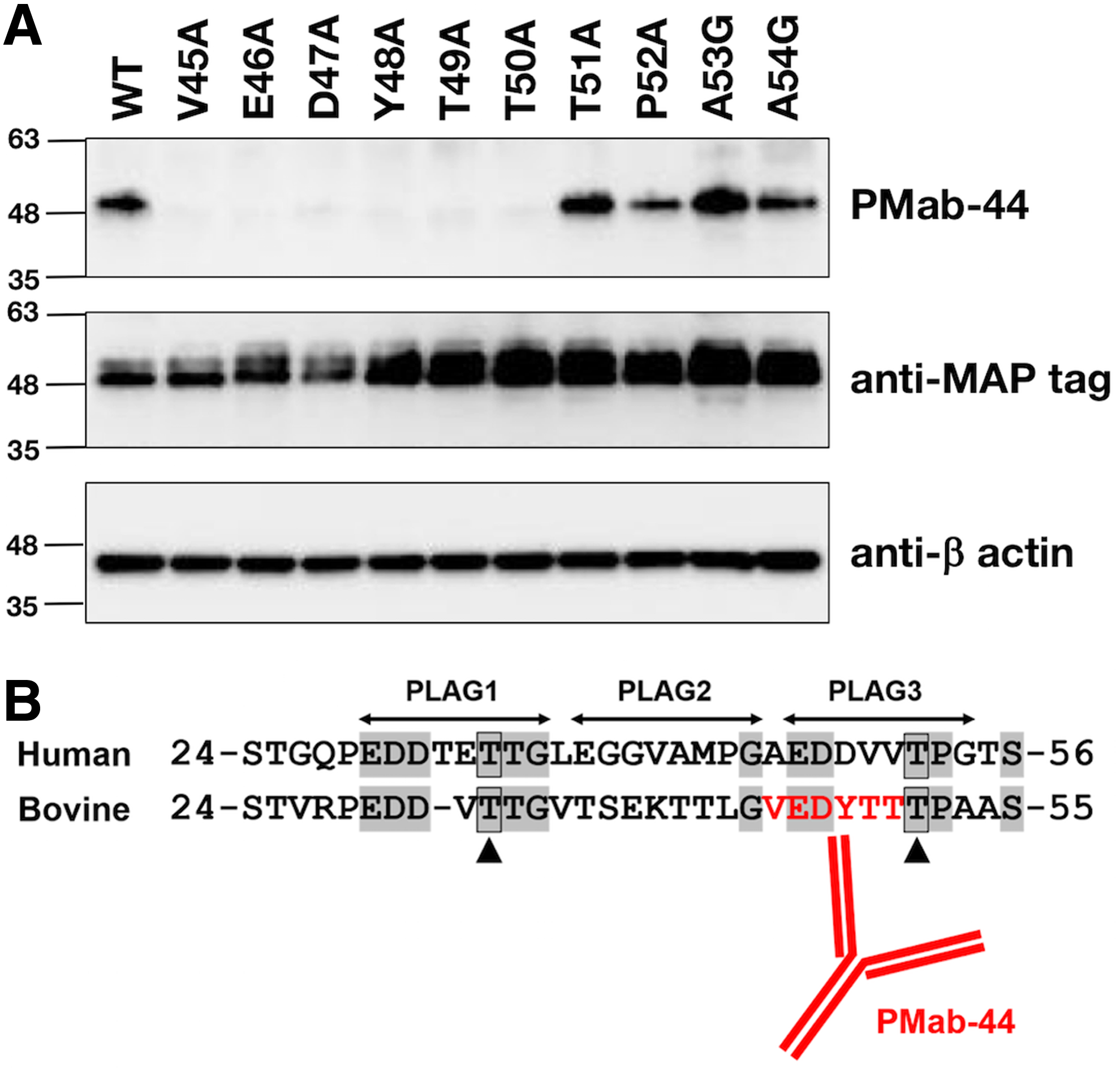
Epitope mapping of an anti-bovPDPN mAb PMab-44 using point mutants by western blot analysis.
Determination of the binding affinity using flow cytometry
We next performed a kinetic analysis of the interaction of PMab-44 with CHO/bovPDPN using flow cytometry. As shown in Figure 4, the KD of PMab-44 was determined as 3.3 × 10−9 M, indicating that PMab-44 possesses a high affinity against bovPDPN.
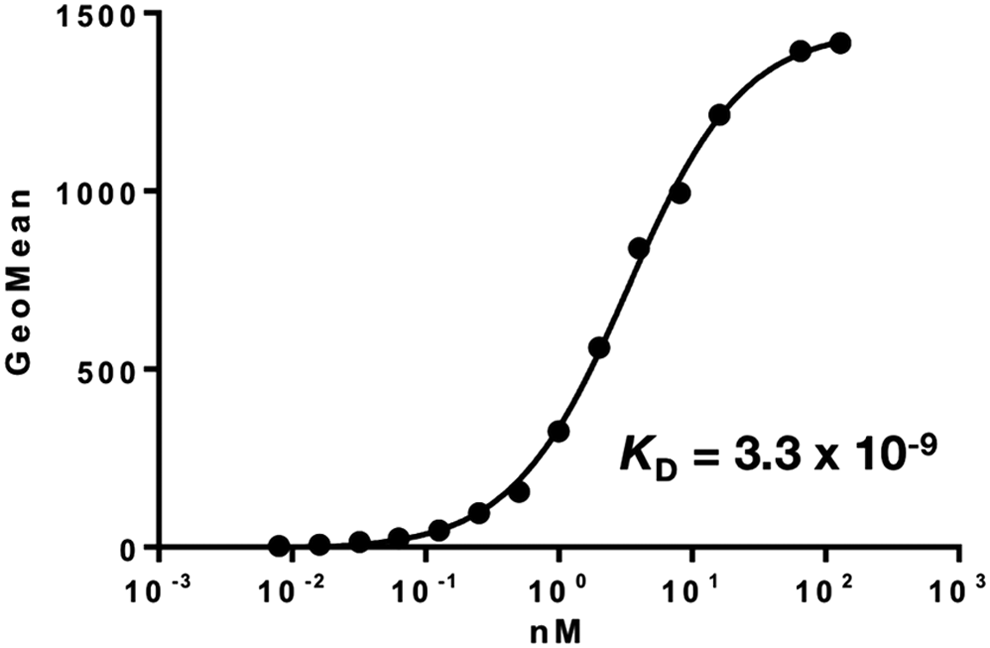
Determination of binding affinity using flow cytometry. CHO/bovPDPN cells were resuspended with 100 μL of serially diluted antibodies (0.6 ng/mL–10 μg/mL) followed by secondary anti-mouse IgG. Fluorescence data were collected using a cell analyzer. CHO, Chinese hamster ovary.
In conclusion, the most important amino acids for PMab-44 recognition against bovPDPN were determined to be Glu46, Tyr48, and Thr50; second in importance were Val45, Asp47, and Thr49. Three amino acids, Glu46, Tyr48, and Thr50, exist in the PLAG3 of bovPDPN,(25) indicating that the PLAG3 of bovPDPN is also important for immunogenicity in producing useful anti-PDPN mAbs.
Footnotes
Acknowledgments
This research was supported in part by grants from the Science and Technology Research Promotion Program for Agriculture, Forestry, Fisheries, and Food Industry, Japan (grant 26058B to S.K.), and the NARO, Bio-oriented Technology Research Advancement Institution (the special scheme project on regional developing strategy: grant 16817557 to S.K.). This work was also supported in part by Project for utilizing glycans in the development of innovative drug discovery technologies from Japan Agency for Medical Research and Development, AMED (Y.K.), by the Basic Science and Platform Technology Program for Innovative Biological Medicine from AMED (Y.K.), by the Platform for Drug Discovery, Informatics, and Structural Life Science (PDIS) from AMED (Y.K.), by the Regional Innovation Strategy Support Program from the Ministry of Education, Culture, Sports, Science and Technology (MEXT) of Japan (Y.K.), by JSPS KAKENHI Grant Number 26440019 and 17K07299 (M.K.K.) and 16K10748 (Y.K.). This work was performed in part under the Cooperative Research Program of Institute for Protein Research, Osaka University, CR-16-05 and CR-17-05 and by the Grant for Joint Research Project of the Institute of Medical Science, the University of Tokyo. The authors would like to thank Enago (
Author Disclosure Statement
No competing financial interests exist.