
Abstract
Type II diabetes mellitus (T2DM) is the prevalent type of diabetes, including 90% of the cases world-wide. Helicobacter pylori plays a pathogenic role in the development of T2DM. The host genetic factors have a significant impact on the clinical outcome and anatomical distribution of H. pylori infection and polymorphisms in several genes such as tumor necrotic factor (TNF)-α and mannose-binding lectin (MBL) and are considered to increase the risk for the development of T2DM. In this study, we investigate the prevalence rate of H. pylori infection and its relationship to MBL rs1800450 and TNF-α rs1800620 polymorphism in T2DM. In this case–control study, 174 patients with type II diabetes and 185 healthy controls were studied. Also, demographics, physical, and biochemical parameters were performed in all patients. The DNA extracted from blood specimens was amplified by H. pylori cagA-specific primers. The MBL rs1800450 and TNF-α rs1800620 genotyping were detected by amplification refractory mutation system–polymerase chain reaction (ARMS–PCR). The results show that H. pylori cagA positivity was detected in 42.82% of the diabetic patients and in 22.16% of the control group, and H. pylori infection was closely correlated with MBL rs1800450 AA genotype and TNF-α rs1800620 GG genotype when compared with healthy controls. Furthermore, these two genotypes were strongly associated with H. pylori cagA(+) samples when compared with cagA(−) samples. In addition, the presence of H. pylori cagA(+) infection was significantly associated with the elevated serum levels of total cholesterol and low-density lipoprotein cholesterol. In general, it can be concluded that molecular analysis of MBL rs1800450 AA genotype and TNF-α rs1800620 AA genotype is important in the early detection and treatment of T2DM with H. pylori cagA(+) infection.
Introduction
T
Moreover, T2DM is the prevalent type of diabetes, including 90% of cases in the world and Helicobacter pylori plays a pathogenic role in the development of T2DM. This bacteria is a gram-negative, spiral-shaped, microaerophilic bacterium that colonizes more than half of the world population, causing chronic infection, and more prevalent in developing rather than the developed countries.(4,5)
The H. pylori eradication improves dyslipidemia and insulin resistance and decreases inflammation. Severity of clinical outcomes by H. pylori associates by several factors such as cagA, vacA, and dupA. However, these factors alone are not sufficient to explain why so few individuals harboring H. pylori develop T2DM and gastric disease.(6–8)
Tumor necrotic factor (TNF) as chemotactic factors is present in gastric epithelium, and it can induce a number of changes in the gastric epithelium that promote inflammation and epithelial damage, thus leading to increased risk of aberrant repair giving the picture of gastric atrophy or epithelial cell metaplasia.(9,10) Several studies have reported a relationship between H. pylori infection and an increase in the inflammatory gene response characterized by the upregulation of several genes such as interleukin (IL)-1β, IL-1RN, IL-8, IL-10, and TNF-α. However, the TNF-α is a very important multifunctional proinflammatory cytokine and provides a rapid form of host defense against infection by the H. pylori. The host genetic factors have a significant impact on the clinical outcome and anatomical distribution of H. pylori infection, and polymorphisms in TNF-α are considered to increase the risk for the development of T2DM.(11,12) Several polymorphisms in the promoter region of the TNF-α gene have been associated with a variety of inflammatory diseases and cancer. TNF-α rs1800620 (−308 G >A), one of the most frequently studied polymorphisms on this gene, seems to have an influence on transcriptional activities and may contribute to the development of T2DM. The H. pylori infection can induce an inflammatory response that may be influenced by rs1800620 polymorphisms and gene expression of the TNF-α gene.(13,14)
Furthermore, TNF-α has an important role in host defense against H. pylori infection, but a high concentration of this cytokine may cause severe pathology. TNF-α production is regulated, in part, at the transcriptional level, and many studies have implicated TNF-α polymorphisms as potential determinants of T2DM susceptibility.(15–17)
On the contrary, mannose-binding lectin (MBL) is a C-type lectin secreted by the liver and is a component of the innate immune defense system. Recent studies have further implicated MBL deficiency in the development of T2DM, type 1 diabetes, gestational diabetes mellitus, diabetic nephropathy, insulin resistance, and obesity.(18–20) In addition, the MBL deficiency is associated with autoimmune, inflammatory, infectious, and vascular disease. Serum MBL levels in T2DM strongly associate with the presence of several polymorphisms within the MBL2 gene such as MBL rs1800450 (codon 54: G >A).(21–25) Genotyping of bacterium and host gene polymorphisms could be important in the early identification of individuals at high risk of developing T2DM. Therefore, the aim of the present study was to evaluate the prevalence rate of H. pylori infection in T2DM and its relationship with MBL rs1800450 and TNF-α rs1800620 polymorphism.
Materials and Methods
Physical and biochemical parameter
In this case–control study, 174 patients with T2DM and 185 healthy controls were studied. Also, demographics of the patients such as age, gender, and duration of disease were documented. Height and weight were measured and body mass index (BMI) was calculated as weight/height 2 (kg/m2). Waist–hip ratio (WHR) was calculated as waist circumference divided by hip circumference. Homeostatic model assessment (HOMA) method, which has been validated as a reliable measure of insulin sensitivity in vivo in humans, was used to estimate insulin resistance (HOMA-IR). Systolic and diastolic blood pressure (SBP and DBP) measurements using standard sphygmomanometry were performed. Biochemical investigation, including hemoglobin A1c (HbA1c), cholesterol, triglyceride, high-density lipoprotein (HDL), low-density lipoprotein (LDL), and fasting blood glucose, was performed in all patients.
DNA extraction and H. pylori detection
CINAGEN, Inc.'s DNA Extraction Kit was used to isolate blood. The extraction procedure was performed according to the manufacturer's instruction for blood samples. The DNA from blood specimens was amplified by H. pylori cagA primers. The PCR-primers are described in Table 1. For detection cagA of H. pylori, PCR amplification was performed using two primer pairs of cagA-FW and cagA-RW.
F, forward; MBL, mannose-binding lectin; R, reverse.
MBL rs1800450 genotyping by PCR-restriction fragment length polymorphism
The detection of MBL rs1800450 genotypes was performed by PCR-restriction fragment length polymorphism analysis. For MBL rs1800450 genotyping, in addition to PCR, A and G alleles were detected by Ban I restriction enzyme digestions of the 328-bp product amplified by the allele P (Table 1), followed by a 2.5% agarose gel electrophoresis. Ban I cleaves the G allele into two fragments allele (245 and 83 bp) and leaves the A allele undigested.(22)
PCR amplification was performed using 100 ng of DNA derived from samples with primers and PCR amplification conditions.
All PCRs were initiated by a 10-minute denaturizing step at 94°C and completed by a 7-minute extension step at 72°C. The temperature cycles for different types of PCRs were as follows: 32 cycles of 40 seconds at 94°C, annealing temperature for 40 seconds and 72°C for 55 seconds (PaqLab, Germany). The PCR fragments were run in 2% agarose gel and were visualized by SYBR Green staining.
TNF-α rs1800620 genotyping by ARMS-PCR
The detection of TNF-α polymorphism at positions −308 was performed by ARMS-PCR analysis. For each TNF-α -308 G/A genotypes, three primers (one common, one specific for the mutant, and one specific for the wild-type allele) were used.(24) The PCR-primers are described in Table 2.
PCR amplification was performed using 100 ng of DNA derived from samples with primers and PCR amplification conditions. The reaction mixture underwent initial denaturation process at 94°C for 5 minutes, followed by 35 cycles at 94°C for 60 seconds, 55°C for 60 seconds, and 72°C for 35 seconds. The final extension was performed at 72°C for 10 minutes in thermocycler. The PCR fragments were run in 2% agarose gel and were visualized by SYBR Green staining.
Statistical analyses
Epi Info package program was used for statistical analysis of data. Associations between genotypes, clinical diseases, and virulence markers were evaluated. Statistical analyses were performed by χ2 and p-values <0.05 were considered statistically significant.
Results
Comparison physical parameters and biochemical parameters in T2DM and control
The relationship between patient and control groups by physical and biochemical parameters are shown in Tables 3 and 4. There were differences in WHR, BMI, SBP, or DBP distribution between the study groups, but no differences in gender or age distribution.
BMI, body mass index; DBP, diastolic blood pressure; SBP, systolic blood pressure; WHR, waist–hip ratio.
HbA1c, hemoglobin A1c; HDL, high-density lipoprotein; HOMA, homeostatic model assessment; IR, insulin resistance; LDL, low-density lipoprotein.
In other sides, the patients were significantly associated with the elevated serum levels of total cholesterol, LDL cholesterol, triglyceride, insulin, glucose, HbA1c, and HOMA-IR, but were not associated with HDL cholesterol levels.
Results of cagA detection by PCR
Prevalence of H. pylori infection was significantly higher in diabetes compared with controls. In patients, H. pylori cagA positivity was found in 78 (44.82%), while it was present in only 41 (22.16%) of controls, which was found to be significant (χ2 = 21.28, p = 0.00001) (Fig. 1).
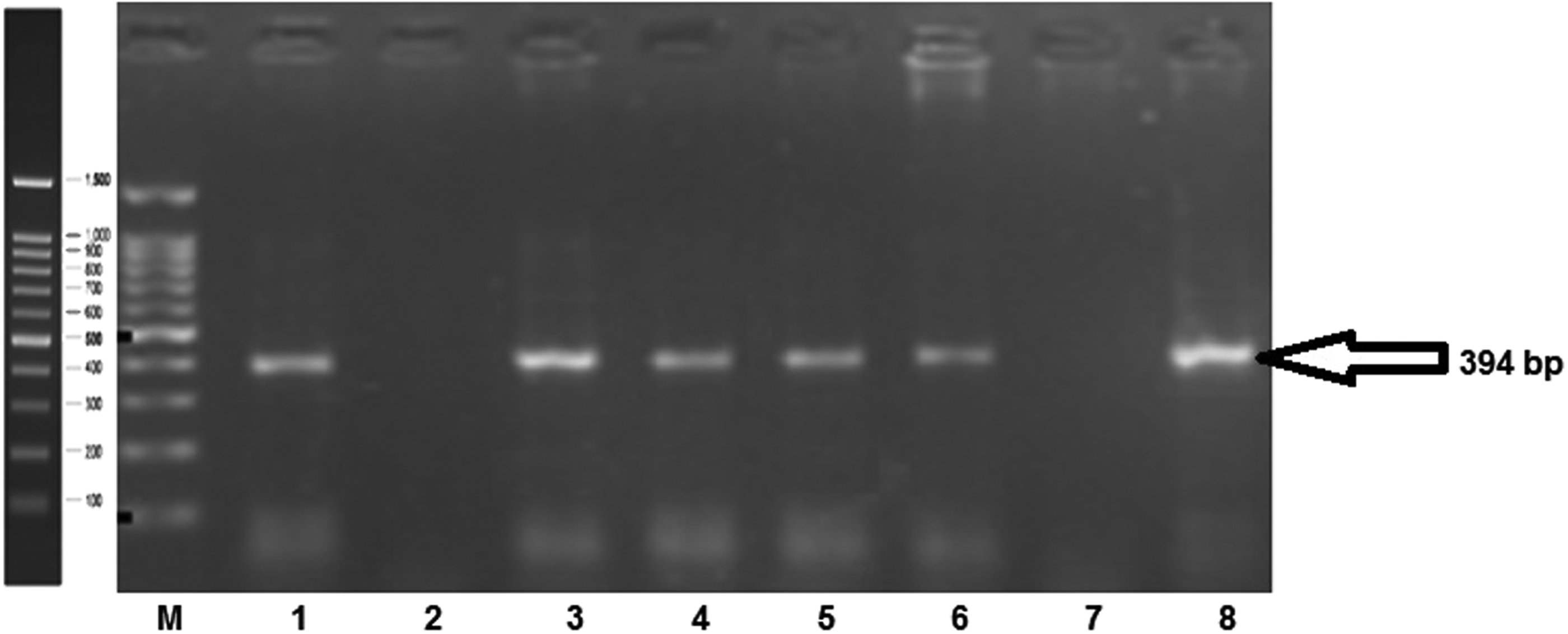
Electrophoresogram of PCR products, Lane M, 100-bp ladder; lanes 1, 3, 4, 5, 6, and 8, samples Helicobacter pylori cagA positive. PCR, polymerase chain reaction.
MBL rs1800450 genotyping
On another side, the frequency of MBL rs1800450 AA genotyping among diabetics with H. pylori infection was 19% and without H. pylori infection was 5% (χ2 = 7.71, p = 0.005) (Table 5; Fig. 2).
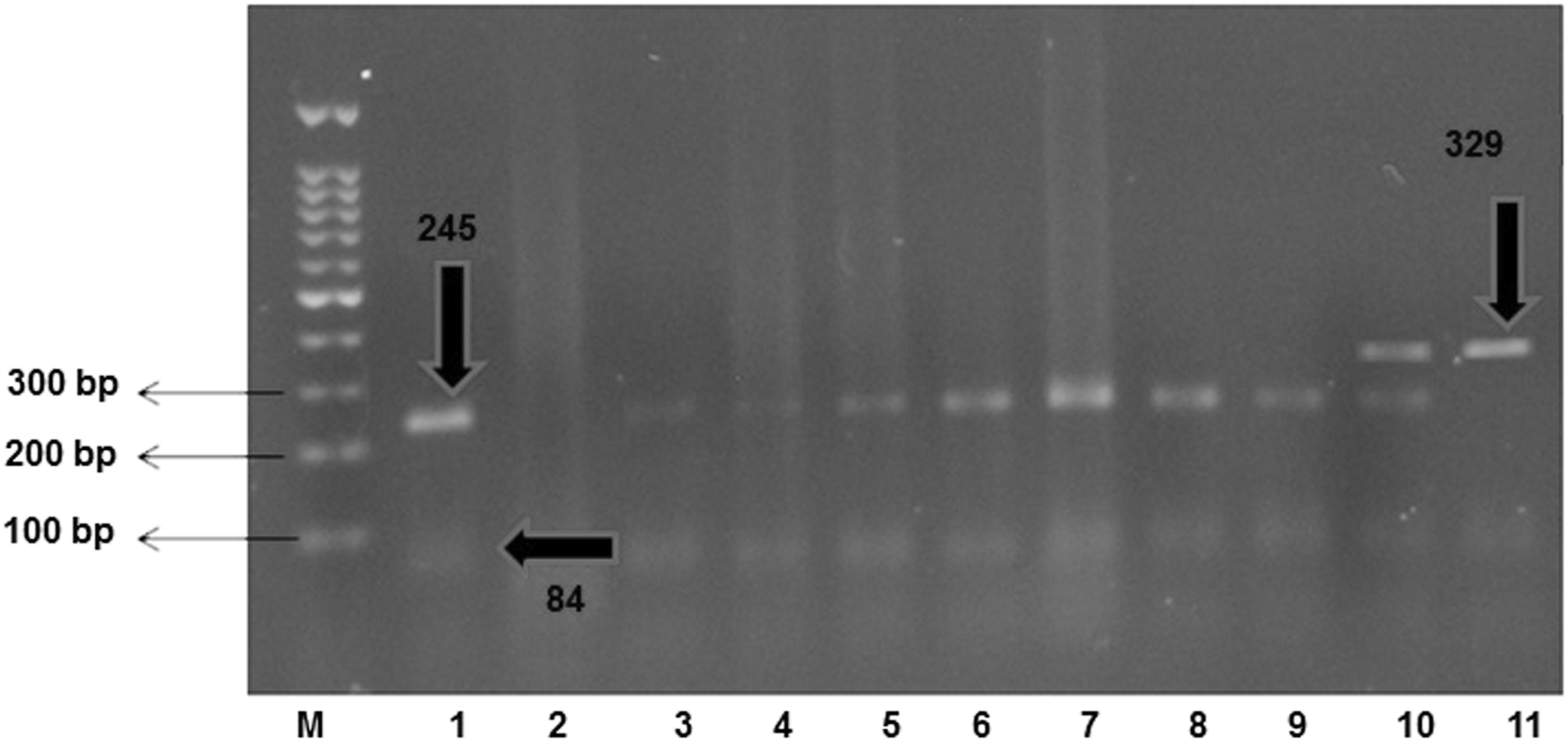
Electrophoresogram of PCR-RFLP products, Lane M, 100-bp ladder. RFLP, restriction fragment length polymorphism.
χ2 = 7.71, p = 0.005.
OR, odds ratio.
The frequency of MBL rs1800450 genotyping in diabetic patients with H. pylori cagA(+) and H. pylori cagA(−) are shown in Table 6.
χ2 = 8.86, p < 0.003.
TNF, tumor necrotic factor.
TNF-α rs1800620 genotyping
The genotype frequencies of the TNF-α rs1800620 studied are shown in Table 5. Interestingly, the frequency of AA genotype was significantly higher in diabetes compared with controls. The frequency of TNF-α rs1800620 AA genotype among diabetic patients was 28.16% and in control was 22.16% (χ2 = 8.86, p < 0.003) (Table 6).
In addition, a significant relationship between TNF-α rs1800620 AA genotypes and the presence of H. pylori was detected. Individuals carrying the AA homozygous genotype have a 3-times greater risk of presenting with T2DM than an individual with the GG homozygous genotype.
In general, it can be concluded that the MBL rs1800450 AA genotype and TNF-α rs1800620 AA genotype were strongly associated with H. pylori cagA(+) samples when compared with cagA(−) samples. In addition, the presence of H. pylori cagA(+) infection was significantly associated with the elevated serum levels of total cholesterol and LDL cholesterol. Furthermore, molecular analysis of MBL rs1800450 AA genotype and TNF-α rs1800620 AA genotype are important in the early detection and treatment of T2DM with H. pylori cagA(+) infection.
Discussion
Patients with T2DM are often affected by chronic infections and H. pylori detects as a public health problem worldwide. Moreover, the T2DM pathogenesis is a complex process resulting from interaction between genetic and environmental factors. Several factors are correlated with the severity of clinical outcomes such as the H. pylori cagA and host genetic factors. The cytotoxin-associated gene (cagA) is a virulence factor that has been well established as being associated with the development of peptic ulceration or gastric adenocarcinoma by H. pylori.(1–8)
While the findings of studies on the prevalence of H. pylori in T2DM patients are contradictory, in our study, H. pylori positivity was detected in 42.82% of the T2DM patients and in 22.16% of the control group. While the prevalence of H. pylori infection was 74.4% in T2DM patients and 50% in the control group, the prevalence of H. pylori was significantly higher in the T2DM group compared with the control group.(16) The presence of cagA gene has been associated with higher grades of inflammation, which may lead to the development of T2DM. In our study, patients with T2DM had significantly higher values for BMI, triglyceride, HbA1c, glucose, total cholesterol, WHR, LDL cholesterol, HOMA-IR, SBP and DBP and insulin, and lower levels of HDL cholesterol than control participants (all p < 0.0001). However, some studies did not find any significant difference in the T2DM group and the control group with regard to H. pylori infections. It was detected that H. pylori infection significantly increases insulin resistance in asymptomatic patients.(17) In another study, the prevalence of macrovascular (cardiovascular and cerebrovascular diseases) and microvascular (nephropathy, neuropathy, and retinopathy) complications is significantly higher in H. pylori-positive diabetic patients.(4) Furthermore, there are studies which report that microalbuminuria is significantly higher in H. pylori-positive patients, regardless of the development of diabetes. Also, there was a significant relationship between H. pylori-positivity and neuropathy.(5)
However, bacterial factors or host genetic factors alone are not sufficient to explain the diverse outcomes of H. pylori-related diseases. The strong association between the inflammatory process and T2DM pathogenesis is very important in the evaluation of genetic polymorphisms in host genetic factors such as MBL and TNF-α genes.(16–19)
The MBL is a component of the innate immune defense system that was secreted by the liver, and MBL deficiency is associated with autoimmune, inflammatory, infectious, and vascular disease.(20–22) The polymorphism in the MBL gene may influence not only gene expression but also susceptibility to disease and pathogenesis. Furthermore, the relationship among genes, host polymorphisms, their expression, and the severity of clinical outcomes in the ethnically diverse Iranian population needs to be further characterized.(18–25) We believe that the association between the host genetic factors and the genetic factors of H. pylori can determine the severity of T2DM disease in infected individuals.
It was shown that the MBL rs1800450 and MBL rs11003125 polymorphism contributed to T2DM susceptibility in both Native Americans and Old Order Amish.(18) Association was detected between MBL rs1800450 and T2DM and diabetic nephropathy and the influence of MBL2 polymorphisms on serum MBL levels.(19)
The observations of the present study reveal that rs1800450 AA genotypes and H. pylori cagA are associated with increased susceptibility to T2DM. This evidence suggested that the AA genotype of rs1800450 may be harmful to patients with T2DM with H. pylori cagA(+) infection.
The human MBL is a 32 kDa protein consisting of 248 amino acids encoded by MBL2 mapped to 10q21.1 and is an important component of the innate immune system.(18–21) The MBL plays a significant role in the defense against various infections and recognizes the carbohydrate domain on the microbial surface, opsonizes the pathogen, and/or activates lectin pathways. In another study, an association was shown between rs1800450 and susceptibility to severe malaria.(20) Furthermore, MBL2 rs1800450 has been associated with various diseases such as filariasis, leishmaniasis, tuberculosis, HIV infection, and rheumatoid arthritis.(21–25)
On the contrary, TNF-α inhibits insulin transduction, has an effect on glucose metabolism, plays a role in the development of obesity, insulin resistance, and the possibility of T2DM, and is involved in inflammatory responses.(9–16) TNF-α rs1800620 polymorphism in different ethnic populations was investigated and detected a weak association between the A variant and gastric cancer risk in Caucasian populations. In this study, no associations were found between this A variant and gastric carcinogenesis in East Asian samples or other ethnic populations.(9) It was demonstrated that patients with chronic gastritis infected by cagA-positive strains of H. pylori had the highest cytokine messenger RNA (mRNA) and protein levels.(10) Santos et al. found that TNF-α rs1800620 polymorphism apparently does not affect the presence of H. pylori or the progression of gastric disease.(11)
The TNF-α gene is located within the HLA III region in chromosome 6p21 and involved in inflammatory responses. This gene encodes a potent cytokine that has been implicated as an important factor in obesity-associated insulin resistance and pathogenesis of T2DM. In another study, it was detected high mRNA expression of IL-6, IL-10, IFN-γ, and TNF-α from H. pylori-infected children and uninfected children in the gastric mucosa.(12) However, H. pylori cagA-positive strains were associated only with higher IL-6 and IL-10 mRNA expression in relationship to the cagA-negative strains. Meanwhile, TNF-α presented increased protein levels and was correlated with bacterial density. It was determined that the AA homozygous genotype presents a greater capacity for producing TNF-α cytokines than other genotypes.(13) In our study, we found a significant difference in the distribution of AA genotype between the study groups. But another study reported that the A variant of the TNF-α-308 polymorphism presented higher transcriptional activity and increased TNF-α cytokine production.(14) Another study found an increased tumor necrosis factor receptor 1 expression in human colorectal adenomas, although TNF-α expression did not differ significantly.(15) In our study, TNF-α rs1800620 AA genotype was statistically significant between patients with H. pylori cagA-positive strains and H. pylori cagA-negative strains, and the evidence was strong enough to reach the conclusion that TNF-α rs1800620 AA genotype was associated with T2DM pathogenesis in H. pylori cagA-positive strains. In other words, it can be concluded that molecular analysis of MBL rs1800450 AA genotype and TNF-α rs1800620 AA genotype is important in the early detection and treatment of T2DM with H. pylori cagA(+) infection.
Footnotes
Acknowledgments
We thank all the patients for their kind collaboration and also the Islamic Azad University for supporting this research. Finally, we thank the head and physicians of the Emam Khomainy Hospital and Medical Genetics Center of Gene Gostar, Karaj, Iran, for help during this research.
Author Disclosure Statement
No competing financial interests exist.