
Abstract
Podoplanin (PDPN), a type I transmembrane sialoglycoprotein, is extensively expressed by normal lymphatic endothelial cells, renal podocytes, and pulmonary type I alveolar cells. Nevertheless, increased expression of PDPN in malignant tumors not only associates with poor prognosis but also facilitates hematogenous metastasis through interaction with C-type lectin-like receptor-2 presented on platelets, followed by PDPN-mediated platelet activation. We previously reported a novel PMab-38 antibody, an anti-dog PDPN (dPDPN) monoclonal antibody, which specifically recognizes PDPN in squamous cell carcinomas melanomas and cancer-associated fibroblasts in canine cancer tissues. However, the specific binding with the epitope of PMab-38 remains undefined. In this study, flow cytometry was utilized to investigate the epitope of PMab-38, which was determined using a series of deletion or point mutants of dPDPN. The results revealed that the critical epitope of PMab-38 is Tyr67 and Glu68 of dPDPN.
Introduction
P
Recently, we developed a novel PMab-38 mAb, which specifically recognizes dPDPN in renal podocytes, in contrast to LECs or type I alveolar cells, by immunohistochemical analysis.(24) Moreover, PMab-38 also recognizes dPDPN of SCCs, melanomas, and cancer-associated fibroblasts (CAFs) in cancer tissues.(12,25) Therefore, this study was initiated to investigate the epitope of PMab-38 using deletion or point mutants of dPDPN in Chinese hamster ovary (CHO)-K1 cells by flow cytometry.
Materials and Methods
Cell lines
CHO-K1 was procured from the American Type Culture Collection (ATCC, Manassas, VA). CHO-K1 cells were transfected with the dPDPN-MAP tag plasmid using Lipofectamine LTX (Thermo Fisher Scientific, Inc., Waltham, MA) according to manufacturer's protocol. CHO-K1 and transfectants were cultured in RPMI 1640 medium (Nacalai Tesque, Inc., Kyoto, Japan) and supplemented with 10% heat-inactivated fetal bovine serum (Thermo Fisher Scientific, Inc.), 100 U/mL of penicillin, 100 μg/mL of streptomycin, and 25 μg/mL of amphotericin B (Nacalai Tesque, Inc.) at 37°C in a humidified atmosphere of 5% CO2 and 95% air. Zeocin (0.5 mg/mL; InvivoGen, San Diego, CA) was added for producing and culturing each transfectant.
Production of dPDPN mutants
The amplified dPDPN complementary DNA was subcloned into a pCAG vector (Wako Pure Chemical Industries, Ltd.) and a MAP tag was added at the N-terminus. Deletion mutants of dPDPN were generated using a HotStar HiFidelity PCR (Qiagen, Inc., Hilden, Germany) with oligonucleotides containing the desired mutations. PCR amplicons bearing the desired mutations were subcloned into pCAG vector using an In-Fusion PCR cloning kit (Clontech, Palo Alto, CA). Substitutions of amino acids (aa) to alanine in the dPDPN sequence were achieved using a QuikChange lightning site-directed mutagenesis kit (Agilent Technologies, Inc., Santa Clara, CA).
Flow cytometry
Cells were harvested after brief exposure to 0.25% trypsin/1 mM ethylenediaminetetraacetic acid (Nacalai Tesque, Inc.). After washing with 0.1% bovine serum albumin in phosphate-buffered saline, the cells were treated with 10 μg/mL of PMab-38 and 5 μg/mL of anti-MAP tag (clone: PMab-1) for 35 minutes at 4°C followed by treatment with Alexa Fluor 488-conjugated anti-mouse IgG (1:1000; Cell Signaling Technology, Inc., Danvers, MA) and Oregon Green-conjugated anti-rat IgG (1:1000; Thermo Fisher Scientific, Inc.), respectively. Fluorescence data were acquired using Cell Analyzer EC800 (Sony, Corp., Tokyo, Japan).
Results and Discussion
Previously, we developed a novel PMab-38 mAb, which exhibited high specificity and sensitivity toward dPDPN and is highly suitable for the detection of renal glomerulus podocytes in immunohistochemical analysis.(24) Interestingly, PMab-38 can be used to detect dPDPN expressed by oral and skin SCCs.(25) CAFs and canine melanoma cells were also recognized by PMab-38.(12,25) Taken together, these findings suggest that the epitope mapping of PMab-38 could provide a promising target for the PDPN-based diagnosis and cancer therapy.
For this study, nine deletion mutants of dPDPN were generated (Fig. 1A). Stable transfections of dPDPN-mutant clones were established in CHO-K1 cells, including dN23 (corresponding to 23–169 aa), dN30 (corresponding to 30–169 aa), dN50 (corresponding to 50–169 aa), dN60 (corresponding to 60–169 aa), dN65 (corresponding to 65–169 aa), dN70 (corresponding to 70–169 aa), dN75 (corresponding to 75–169 aa), dN80 (corresponding to 80–169 aa), and dN110 (corresponding to 110–169 aa). All dPDPN deletion mutants were tagged with N-terminal MAP and therefore were analyzed using flow cytometry for epitope mapping of PMab-38. Flow cytometric analysis revealed that PMab-1 (anti-MAP tag mAb) detected all the dPDPN deletion mutants, including dN23, dN30, dN50, dN60, dN65, dN70, dN75, dN80, and dN110 (Fig. 1B, left). However, a significant loss in the reaction of PMab-38 was observed with dN70, dN75, dN80, and dN110 (Fig. 1B, right). These results indicate that N-terminus of PMab-38 epitope is located between 65th and 70th aa.
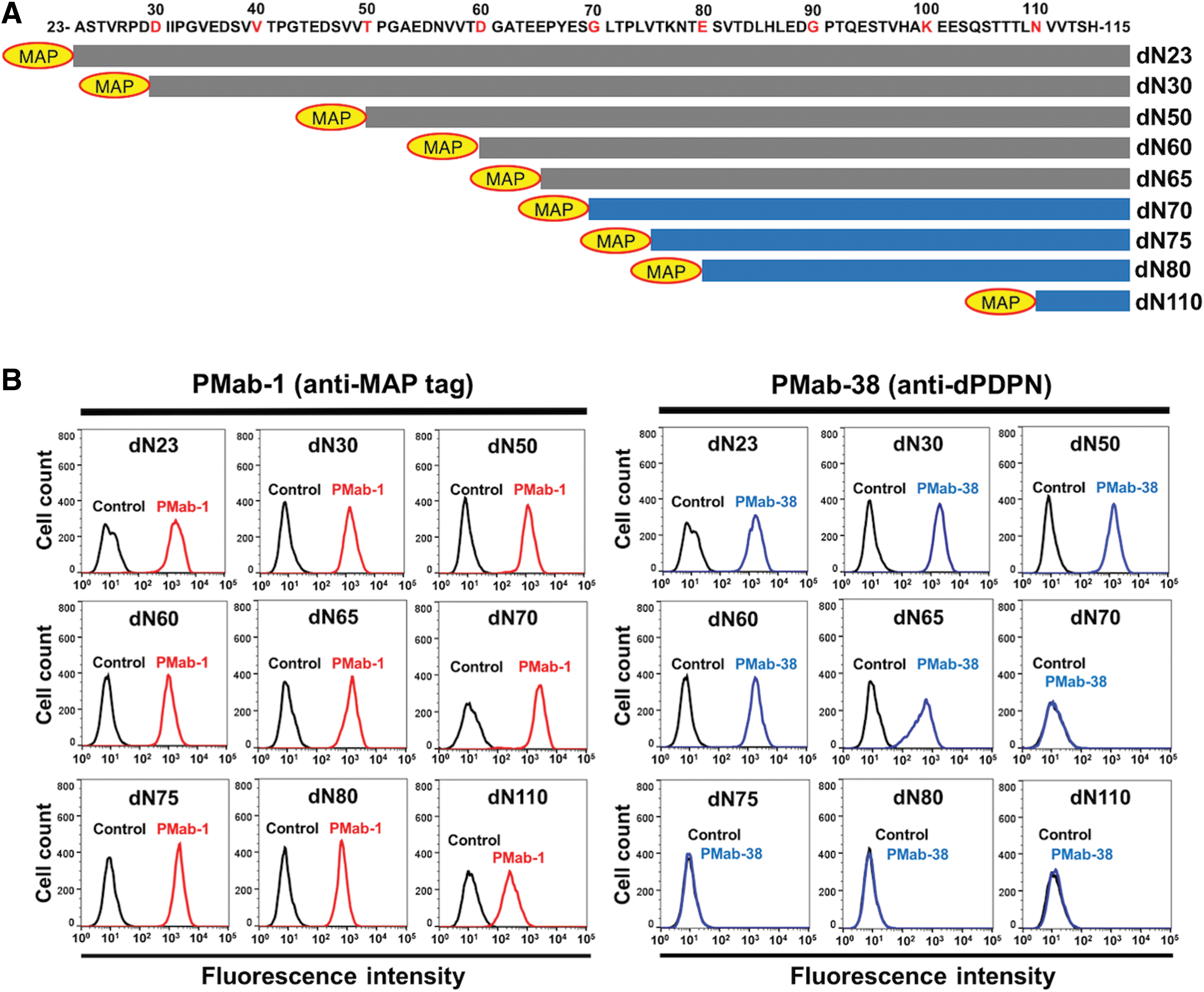
Epitope mapping of PMab-38 using deletion mutants of dPDPN.
Next, to further investigate the crucial epitope of PMab-38, we produced a series of point mutants of dPDPN in CHO-K1 cells, including D60A, G61A, A62G, T63A, E64A, E65A, P66A, Y67A, E68A, S69A, and G70A. All deletion mutants were recognized by PMab-1 (Fig. 2A). Notably, PMab-38 reacted with all these mutants, except with Y67A and E68A (Fig. 2B). These results provide strong direct evidence that Tyr67 and Glu68 are the crucial epitopes of PMab-38 (Fig. 2C). This YE motif of dPDPN is not conserved among the other species.
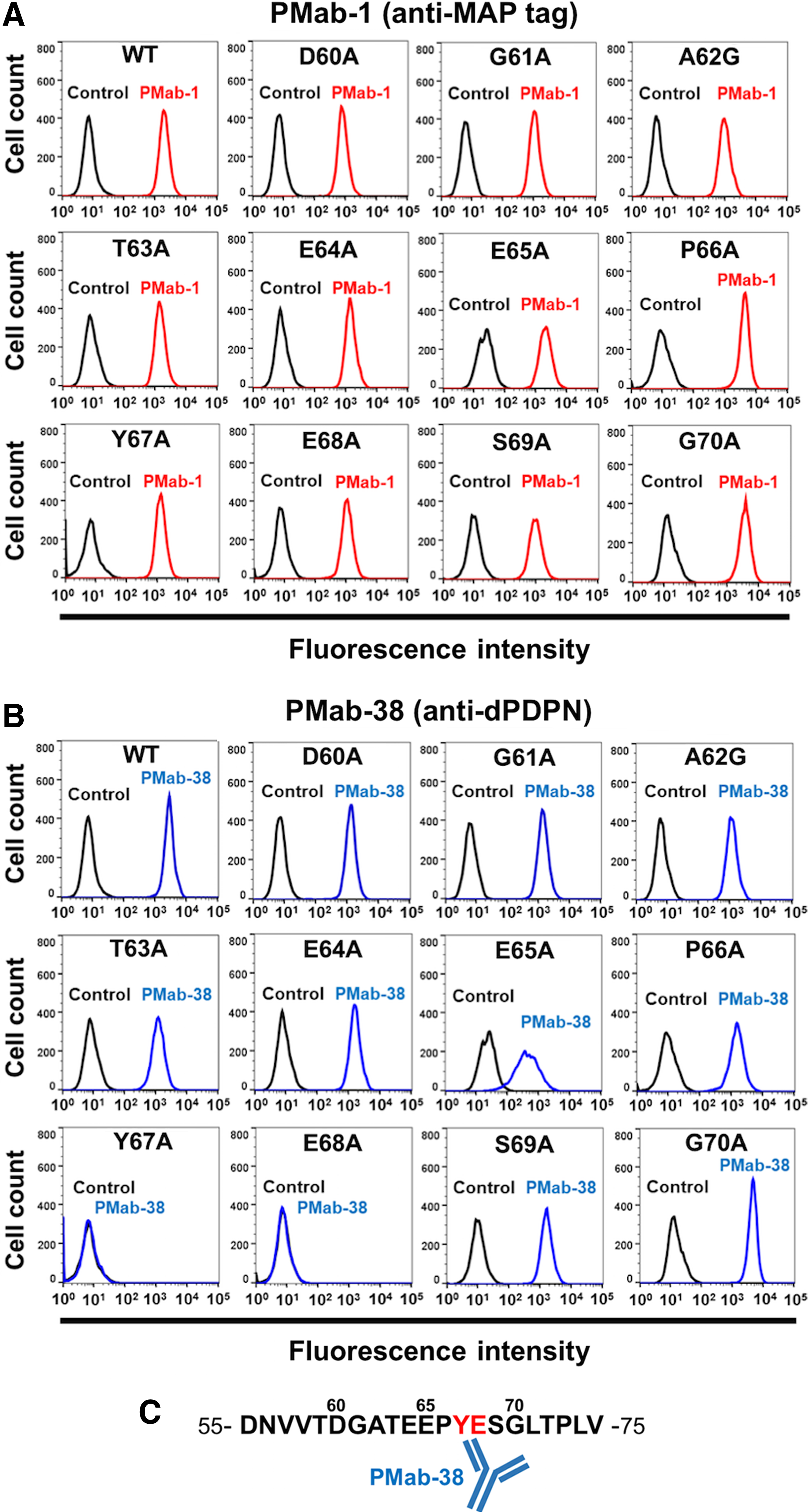
Epitope mapping of PMab-38 using point mutants of dPDPN.
Although detection of PMab-38 for renal glomerulus, SCCs, melanoma, and CAFs using immunohistochemistry and flow cytometry was successful,(12,24,25) our results revealed that PMab-38 has limitation for dPDPN recognition by Western blot analysis (data not shown). One reason might be that PMab-38 has high affinity binding to undenatured dPDPN.
Taken together, these results demonstrated that PMab-38 is a potential diagnostic target for the detection of dPDPN using flow cytometry. Furthermore, the study also revealed Tyr67 and Glu68 as the crucial epitopes of dPDPN for specific binding with PMab-38. This study also provides evidence on the potential application of PMab-38 in unveiling pathophysiological function of dPDPN.
Footnotes
Acknowledgments
This work was supported, in part, by the Basic Science and Platform Technology Program for Innovative Biological Medicine from Japan Agency for Medical Research and development, AMED (Y.K.), by Project for utilizing glycans in the development of innovative drug discovery technologies from AMED (Y.K.), and by Platform Project for Supporting Drug Discovery and Life Science Research (Basis for Supporting Innovative Drug Discovery and Life Science Research [BINDS]) from AMED.
Author Disclosure Statement
Y.K. received research funding from Ono Pharmaceutical, Co., Ltd.