
Abstract
Tumor-targeted antibody–cytokine fusion proteins, called immunocytokines, are expected to be a useful platform for the development of effective antitumor therapeutic agents; however, their design and cost-efficient production remain as challenges. In this study, we constructed an antibody–cytokine fusion protein (Ia1-TNFα) comprising the single-domain antibody Ia1, which targets epidermal growth factor receptor (EGFR) overexpressed in epithelial tumors and a tumor necrosis factor α (TNFα) domain, which has antitumor activity. Ia1-TNFα was produced in a soluble form by using an Escherichia coli expression system, and after affinity purification of the culture supernatant, an yield of ∼2 mg/L of cell culture was obtained. Gel filtration analysis showed that Ia1-TNFα existed predominantly as a trimer, which is consistent with the multimerization state of TNFα. Ia1-TNFα exhibited approximately 7-fold lower TNFα biological activity than that of TNFα itself. Flow cytometric analysis revealed that Ia1-TNFα specifically bound to EGFR-expressing tumor cells and that its binding activity was higher than that of monovalent Ia1, suggesting that the fusion protein bound to the tumor cells multivalently. Altogether, these results show that fusion of TNFα with a single-domain antibody could be a cost-efficient means of producing antitumor therapeutic agents.
Introduction
C
Advances in recombinant DNA technology now allow the construction of antibody–cytokine fusion proteins called immunocytokines. The purpose of fusing a cytokine to an antibody is to harness the targeting capability of the antibody moiety to deliver the cytokine to the site of disease.(4) To date, several immunocytokines with various molecular architectures have been developed, some of which have been evaluated in clinical trials.(5) Generally, the molecular architectures of immunocytokines can be divided into two basic categories: IgG-based immunocytokines that comprise a cytokine fused to an IgG, most often at the N- or C-terminus of its heavy or light chain through a polypeptide linker,(6) and antibody fragment-based immunocytokines, where a cytokine is fused to an antibody fragment (e.g., an antigen-binding fragment or single-chain variable fragment [scFv]) or engineered variant (e.g., a diabody).(7–9) However, one limitation in the development of antibody-based drugs is their poor productivity, which is often the result of the size and complexity of the drug's molecular architecture.(5) To overcome this problem, most of the immunocytokines developed so far have been produced by using mammalian expression systems.(6,8,9)
Among the large number of antibody fragments and antigen-binding scaffold proteins available, single-domain antibodies (sdAbs) are an ideal building block for constructing engineered immunocytokines that have multispecific or multifunctional activities because they are small, highly stable, highly soluble, and are easily genetically modified.(10,11) Recently, we reported that we produced a bispecific antibody tetramer comprising two different sdAbs by using an Escherichia coli expression system.(12)
In the present study, we constructed an antibody–cytokine fusion protein (Ia1-TNFα) specific for epidermal growth factor receptor (EGFR) that contained the sdAb Ia1 and a tumor necrosis factor α (TNFα) domain. TNFα is a TNF superfamily pleiotropic cytokine that forms a 52.5-kDa noncovalently linked homotrimer.(13) TNFα has antitumor activity, and its suitability as an antitumor agent has been investigated in clinical trials.(14) However, its systemic toxicity has limited its use in cancer therapies.(15) Ia1 is an sdAb specific for the receptor tyrosine kinase EGFR, which is a validated target in cancer therapy because it is overexpressed on tumor cells.(16) To examine productivity, we recombinantly produced Ia1-TNFα by using an E. coli expression system and then examined its antitumor capabilities in in vitro experiments.
Materials and Methods
Cells and culture conditions
Human epidermoid carcinoma cell line A431 was obtained from RIKEN Cell Bank (Tsukuba, Japan) and maintained in Dulbecco's modified Eagle's medium supplemented with 10% (v/v) fetal bovine serum. Mouse fibroblast cell lines, NIH3T3 and L929, were kindly provided by Dr. Taro Tachibana (Osaka City University, Japan) and maintained in RPMI 1640 medium supplemented with 10% (v/v) fetal bovine serum. All cells were maintained at 37°C under a humidified atmosphere containing 5% CO2.
Preparation of recombinant proteins
Recombinant Ia1, an sdAb, was expressed in the periplasm of E. coli strain BL21(DE3) by using a previously constructed expression plasmid for C-terminal His6-tagged Ia1(17) and then purified by using immobilized metal ion affinity chromatography with TALON Metal Affinity Resin (Clontech Laboratories, Inc., Mountain View, CA). Recombinant human TNFα was expressed in the cytoplasm of E. coli strain BL21(DE3) by using pET20b vector (Novagen, Madison, WI) containing DNA encoding human TNFα and then purified by a combination of anion exchange chromatography (HiTrap Q XL; GE Healthcare Life Sciences, Tokyo, Japan), gel filtration chromatography (Superdex 75 16/600; GE Healthcare Life Sciences), and cation exchange chromatography (RESOURCE S; GE Healthcare Life Sciences).
Construction of the Ia1-TNFα expression vector
The DNA fragment encoding human TNFα was amplified by means of polymerase chain reaction by using the primers SacII_G1_TNFA (5′-CCCCCGCGGGTGGCGGCGGTAGCGTCCGCAGTTCGAGTCGTAC-3′) and XhoI_FLAG_TNFA (5′-CCCCTCGAGTTTGTCATCATCGTCTTTGTAGTCCAGCGCAATAATGCCGAAG-3′) to add FLAG tag and restriction enzyme sites for SacII and XhoI. The polymerase chain reaction product was digested with SacII and XhoI and then cloned into a modified pET20b vector containing DNA encoding Ia1. The resultant expression vector for Ia1-TNFα was named pET-Ia1-TNFα.
Expression and purification of Ia1-TNFα
E. coli BL21(DE3)pLysS was transformed with pET-Ia1-TNFα and the transformants were grown at 28°C in 2 × YT medium supplemented with 100 μg/mL ampicillin and 34 μg/mL chloramphenicol. When the optical density at 600 nm reached 0.8, isopropyl-β-
Gel filtration analysis
Analytical gel filtration chromatography was performed by using a Superdex 200 10/300 column (GE Healthcare Life Sciences) and an ÄKTA Prime Plus System (GE Healthcare Life Sciences). The column was pre-equilibrated with PBS. Protein samples were injected into the column and eluted with PBS at a flow rate of 0.5 mL/min. Catalase (232 kDa), TNFα (52.5 kDa), chymotrypsinogen (25.0 kDa), and ribonuclease A (13.7 kDa) were used as molecular weight standards.
Flow cytometric analysis for specific binding of Ia1-TNFα
NIH3T3 or A431 cells (5 × 105) were incubated with 2 μg/mL Ia1-TNFα on ice. To inhibit the EGFR-specific binding of Ia1-TNFα, 40 μg/mL Ia1 was also added to the cells. After incubation, the cells were washed with FACS buffer (PBS containing 0.1% NaN3) and then stained with mouse anti-FLAG tag antibody 1E6 (Wako) followed by goat anti-mouse IgG (H+L) antibody–fluorescein isothiocyanate conjugate (KPL, Gaithersburg, MD). Stained cells were analyzed by means of flow cytometry (FACSCalibur; Becton Dickinson, Tokyo, Japan).
EGF competitive binding assay
A431 cells (5 × 105) were incubated with 0.1 μg/mL of biotinylated EGF (Life Technologies, Gaithersburg, MD) in the presence or absence of a competitor (i.e., Ia1 or Ia1-TNFα). After washing the cells with FACS buffer, EGF-bound cells were detected by using streptavidin–Alexa Fluor 488 conjugate (Jackson ImmunoResearch, Westgrove, PA) and counted by using a flow cytometer (FACSCalibur). The percentage of mean fluorescence intensity (% MFI) was calculated with the following equation:
The half-maximal inhibitory concentration (IC50) value was calculated by fitting the data to a sigmoidal dose–response curve by using MasterPlex 2010 software (Hitachi Solutions America, San Francisco, CA).
L929 TNF bioassay
L929 cells were plated into a flat-bottom, 96-well plate at a density of 2 × 104 cells/well and cultured in RPMI 1640 medium supplemented with 10% (v/v) fetal bovine serum. After incubation for 24 hours at 37°C, various dilutions of TNFα or Ia1-TNFα were added in the presence of 0.5 μg/mL actinomycin D and the plates were incubated for a further 18 hours. Cell viability was quantified by using an MTS (3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium inner salt) assay (CellTiter 96 AQueous One Solution Cell Proliferation Assay; Promega, Madison, WI) in accordance with the manufacturer's protocol. The percentage of viable cells was calculated with the following equation:
IC50, which was defined as the quantity of protein that induced 50% lysis of L929 cells, was calculated as described in the section titled “EGF competition assay.”
Results
Construction and preparation of Ia1-TNFα
To preserve the antigen binding and TNFα activities of the two domains in Ia1-TNFα, we fused the Ia1 domain to the N-terminus of the TNFα domain through a flexible Gly4-Ser linker in the same domain order that was used in several previously reported antibody–TNFα fusion proteins (Fig. 1A, B).(18,19)
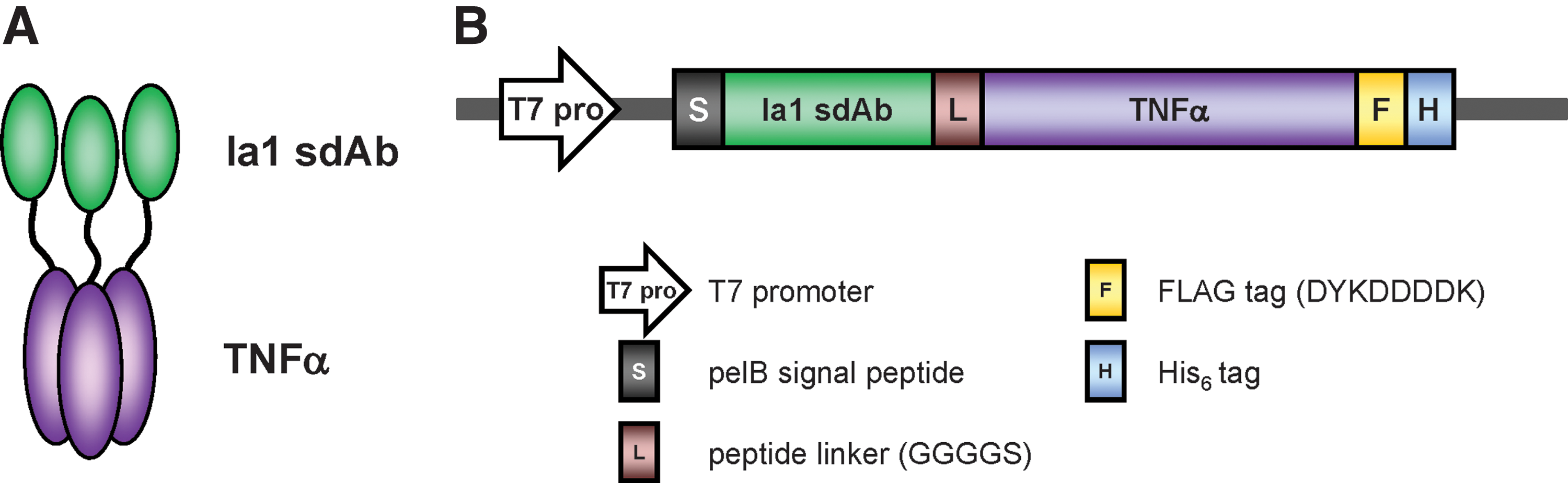
Construction of the fusion protein Ia1-TNFα.
To produce Ia1-TNFα, we used an E. coli expression system. However, we predicted that correct disulfide bond formation, which is critical for correct antibody folding, would not occur under the reducing conditions of the E. coli cytoplasm. Therefore, because disulfide bond formation is catalyzed in the oxidative periplasmic compartment of E. coli,(20) the pelB signal peptide was fused at the N-terminus of Ia1-TNFα to ensure its expression in the bacterial periplasm, rather than the cytoplasm.
Sodium dodecyl sulfate–polyacrylamide gel electrophoresis and western blotting analysis showed that Ia1-TNFα was present in the cell culture supernatant in a soluble form, although it was also detected in an insoluble form (data not shown). Thus, we purified Ia1-TNFα from the cell culture supernatant by using immobilized metal ion affinity chromatography and obtained about 2 mg of pure recombinant protein per liter of cell culture (Fig. 2A).
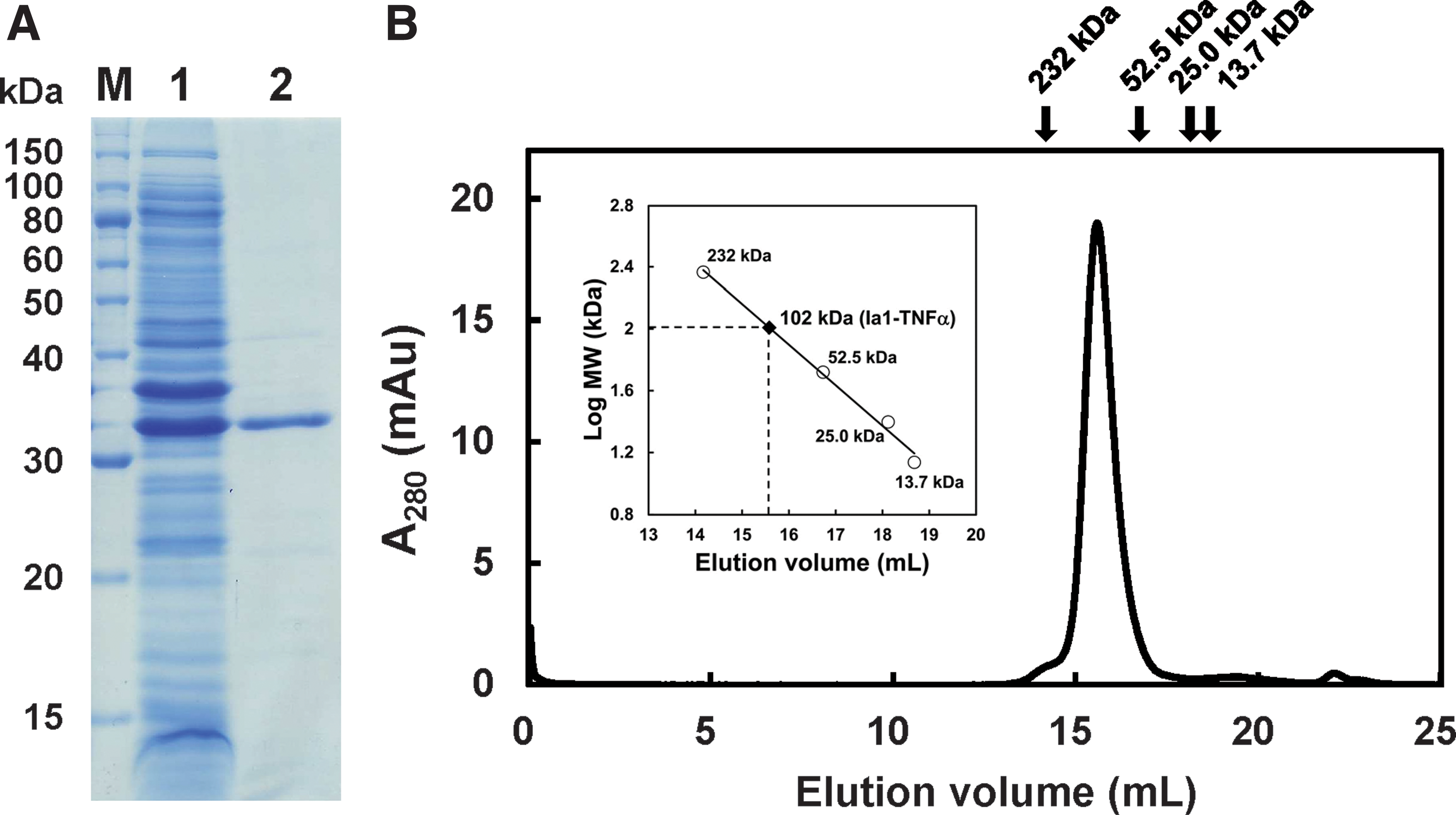
Multimerization states of Ia1-TNFα
To investigate the multimerization states of Ia1-TNFα, we performed gel filtration analysis (Fig. 2B). Ia1-TNFα was eluted as a single peak at a molecular mass of ∼102 kDa, which corresponded with the trimeric molecular weight calculated from the amino acid sequence (i.e., 102 kDa). We previously reported that Ia1 existed as a monomer(12) and TNFα is known to form a trimer.(13) This suggests that Ia1-TNFα formed a trimer through the TNFα domain in the fusion protein.
Binding specificity, antagonistic activity, and avidity of Ia1-TNFα
We next investigated the EGFR binding specificity of Ia1-TNFα by using an EGFR-expressing cell line, A431 (human epidermoid carcinoma cell line), and a cell line that does not express EGFR, NIH3T3 (mouse fibroblast cell line). Flow cytometric analysis showed that Ia1-TNFα bound to A431 cells (Fig. 3A), but not to NIH3T3 cells (Fig. 3B). The binding of Ia1-TNFα to A431 cells was completely inhibited in the presence of excess Ia1, confirming that Ia1-TNFα bound to EGFR rather than to TNF receptor.

Binding properties of Ia1-TNFα. Binding of Ia1-TNFα to A431 cells or NIH3T3 cells was examined by means of flow cytometry.
It is known that the Ia1 competes with EGF to bind to EGFR.(21) Because Ia1-TNFα contains three Ia1 moieties per trimer, we predicted that the increased avidity resulting from the three antigen-binding sites in the fusion protein would provide the fusion protein with greater antagonistic activity against EGFR compared with Ia1. A431 cells (EGFR-expressing cells) were incubated with biotinylated EGF in the presence of various concentrations of Ia1 or Ia1-TNFα (Fig. 4). We found that like Ia1, Ia1-TNFα competed with EGF to bind to A431 cells. The antagonistic activity of Ia1-TNFα (IC50, 5.9 nM) was approximately four times higher than that of Ia1 (IC50, 23 nM), indicating that Ia1-TNFα bound multivalently to EGFR expressed on the surface of A431 cells.

EGF competition assay. A431 cells were incubated with 0.1 μg/mL biotinylated EGF in the presence or absence of various concentrations of Ia1 (open squares) or Ia1-TNFα (closed circles). Bound cells were detected and counted by using streptavidin–Alexa Fluor 488 and a flow cytometer. Data are presented as mean ± SD of triplicate determinations. % MFI, percentage of mean fluorescence intensity; EGF, epidermal growth factor.
TNFα biological activity of Ia1-TNFα
Finally, we assessed the TNFα biological activity of Ia1-TNFα by using L929 cells, which do not express EGFR. L929 cells were incubated with TNFα or Ia1-TNFα in the presence of actinomycin D, and cell viability was determined by using an MTS assay. Ia1-TNFα and TNFα were both cytotoxic to L929 cells in a dose-dependent manner (Fig. 5). The biological activity of Ia1-TNFα (IC50, 1.1 pM) was approximately seven times lower than that of TNFα (IC50, 0.15 pM). This lower biological activity was probably the result of the Ia1 domain interfering with the binding of the TNFα domain to TNF receptor. 18 Altogether, these results and those of the binding specificity experiment showed that Ia1-TNFα possessed both antigen-binding and TNFα activities.
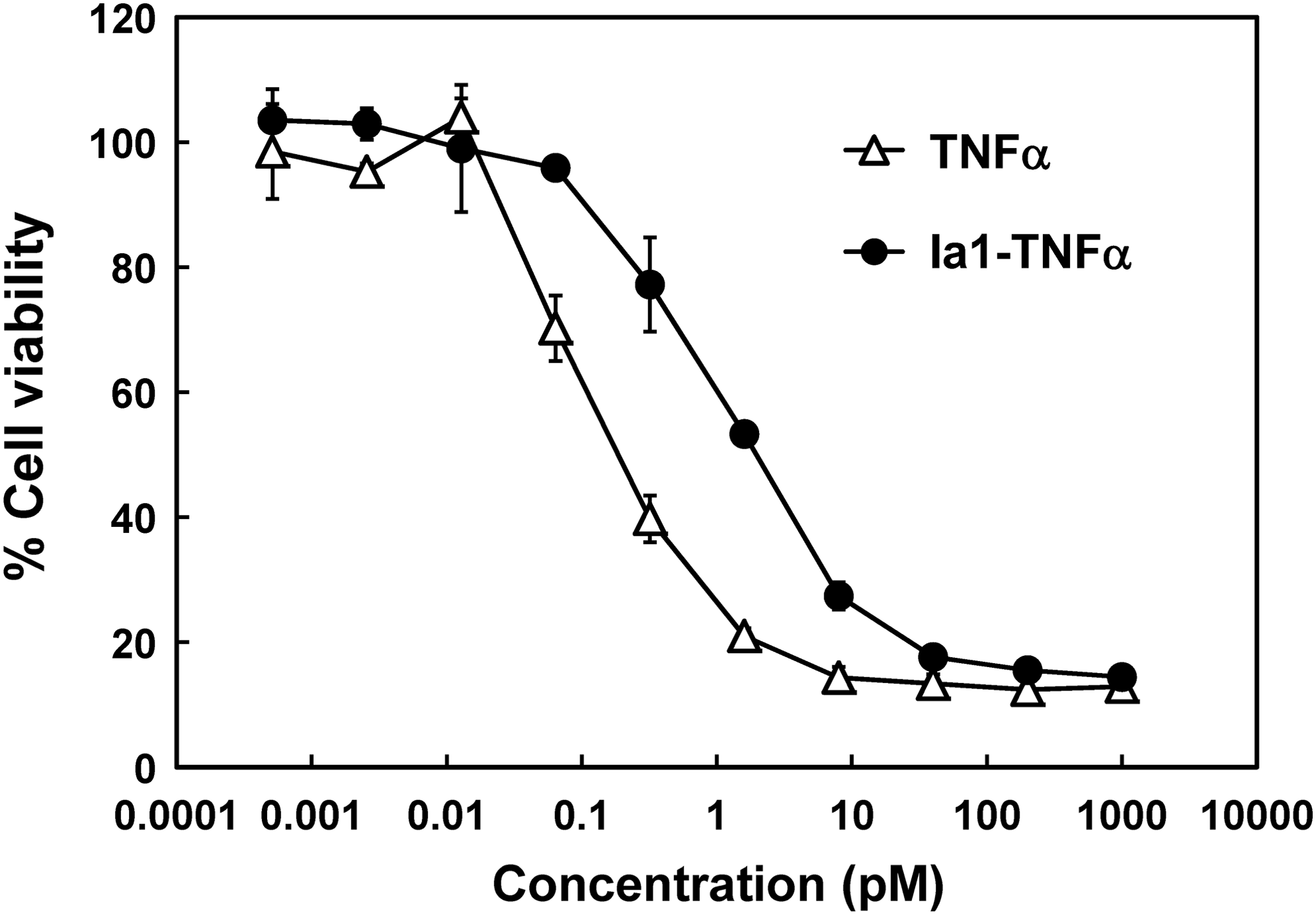
Comparison of the biological activities of TNFα and Ia1-TNFα. L929 cells were incubated with various concentrations of TNFα (open triangles) or Ia1-TNFα (closed circles) for 18 hours in the presence of 0.5 μg/mL actinomycin D. Cell viability was determined by using a 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium inner salt assay. Each symbol and bar represents the mean ± SD of triplicate determinations.
Discussion
In this study, we used an E. coli expression system to produce a novel antibody–cytokine fusion protein specific for EGFR that comprised the sdAb Ia1 and a TNFα domain. A major difficulty in the production of recombinant proteins is that the larger or more complex the protein, the more difficult it is to produce. For example, scFv-based immunocytokines contain more than three domains and therefore are more easily prepared by using expensive mammalian expression systems rather than inexpensive bacterial expression systems.(8,9) Furthermore, some research groups have examined the preparation of antibody–TNFα fusion proteins by refolding insoluble fractions produced by using bacterial expression systems.(18,19) However, it often takes much time and effort to determine the efficient refolding conditions of a recombinant protein. In contrast, the results of the present study suggest that sdAb-based immunocytokines can be easily and inexpensively produced in E. coli.
In the present study, we were able to produce Ia1-TNFα in a soluble form by using a bacterial expression system that preserved the functions of Ia1 and TNFα. Furthermore, we were able to produce ∼2 mg of pure Ia1-TNFα protein from 1 L of E. coli culture. This yield of Ia1-TNFα may have been a result of its simpler structure compared with that of other reported immunocytokines, such as scFv-based immunocytokines, which contain three or more domains. To date, various sdAbs have been used to construct antibody-based fusion proteins because of their small size, high stability, and high solubility.(10,11) However, to our knowledge, the only sdAb-based immunocytokine reported was produced by using a mammalian expression system.(22) The use of sdAb-based fusion proteins is expected to be an advantageous platform for the construction of immunocytokines containing TNF superfamily proteins such as Fas ligand and TNF-related apoptosis-inducing ligand (TRAIL).(23,24) In this context, the results of the present study suggest that bacterial expression systems will be able to be used for cost-efficient production of sdAb-based immunocytokines.
Several other research groups have constructed antibody–TNFα fusion proteins.(25–27) For example, L19-TNFα (Fibromun), an scFv–TNFα fusion protein specific for extra domain B of fibronectin, is a very promising TNFα-based immunocytokine that is currently being examined in Phase I/II clinical trials for its use in the treatment of melanoma.(28,29) However, the in vitro antitumor effect of antibody–TNFα fusion proteins against ErbB family proteins has been modest.(18,19) For example, the EGFR-targeted antibody–TNFα fusion protein generated by Hoffmann et al., scFv(14E1)-TNFα, showed little inhibitory effect on the growth of A431 cells, even in the presence of 5000 U/mL of the protein.(19) Hoffmann et al. suggested that the lack of inhibitory effect on tumor cell growth might have been caused by signals generated upon the binding of the fusion protein to EGFR. Similarly, Ia1-TNFα showed little inhibitory effect on the growth of A431 tumor cells (data not shown). However, a fusion protein composed of lymphotoxin (or TNFβ), which exhibits similar biological activities to TNFα and anti-EGFR antibody 225, has shown in vivo antitumor activity depending on the mouse model used.(30) Therefore, the use of Ia1-TNFα as an anticancer agent warrants further evaluation both in vitro and in vivo.
In conclusion, we constructed a novel antibody–cytokine fusion protein comprising the EGFR-targeted sdAb Ia1 and a TNFα domain. The fusion protein was produced in a soluble form by using an E. coli expression system and it retained the functions of both domains. Thus, the results of the present study suggest that bacterial expression systems are potentially useful for the cost-effective production of immunocytokines targeting EGFR.
Footnotes
Acknowledgments
The authors thank Dr. Taro Tachibana (Osaka City University) for providing the NIH3T3 and L929 cell lines. The human epidermoid carcinoma cell line A431 (RCB0202) was provided by the RIKEN BioResource Center through the National BioResource Project of the Ministry of Education, Culture, Sports, Science, and Technology, Japan. This work was supported by the Japan Society for the Promotion of Science (grant number 25420836).
Author Disclosure Statement
No competing financial interests exist.