
Abstract
Platelets play a vital role in hemostasis and inflammation. The membrane receptor TREM-like transcript-1 (TLT-1) is involved in platelet aggregation, bleeding, and inflammation, and it is localized in the α-granules of platelets. Upon platelet activation, TLT-1 is released from α-granules both in its transmembrane form and as a soluble fragment (sTLT-1). Higher levels of sTLT-1 have been detected in the plasma of patients with acute inflammation or sepsis, suggesting an important role for TLT-1 during inflammation. However, the roles of TLT-1 in hemostasis and inflammation are not well understood. We are developing the mouse model of TLT-1 to mechanistically test clinical associations of TLT-1 in health and disease. To facilitate our studies, monoclonal murine TLT-1 (mTLT-1) antibodies were produced by the immunization of a rabbit using the negatively charged region of the mTLT-1 extracellular domain 122PPVPGPREGEEAEDEK139. In the present study, we demonstrate that two selected clones, 4.6 and 4.8, are suitable for the detection of mTLT-1 by western blot, immunoprecipitation, immunofluorescent staining, flow cytometry and inhibit platelet aggregation in aggregometry assays. In addition, we found that the topical administration of clone 4.8 delayed the wound healing process in an experimental burn model. These results suggest that TLT-1 plays an important role in wound healing and because both clones specifically detect mTLT-1, they are suitable to further develop TLT-1 based models of inflammation and hemostasis in vivo.
Introduction
P
An important platelet membrane protein belongs to the family of receptor proteins known as Triggering Receptors Expressed on Myeloid cells (TREM). The genes for the TREM protein family are clustered on human chromosome 6p21.1 and mouse chromosome 17- 23.99 cM and encode three members of TREM and four of TREM-like transcript (TLT).(3,4) The members of this family are key regulators of innate and adaptive immune response.
The target of our study is the TREM-like transcript-1 (TLT-1), which upon platelet activation is brought to the cell surface to promote thrombus formation and platelet aggregation. During this process a soluble form of TLT-1 (sTLT-1), also contained in the platelet α-granules, is released in the extracellular space, enhancing platelet aggregation in vitro.(5–7) Moreover, sTLT-1 increases platelet reactivity to agonists, suggesting an important role in hemostatic function.(8) These studies provide evidence of aggregation mediated by both forms of TLT-1, making them ideal molecular targets for future studies on thrombotic diseases and inflammatory responses during the hemostatic processes. Murine TLT-1 (mTLT-1) displays a 65% identity to human TLT-1 and is also found in platelet α-granules, and current studies in mice have supported clinical data. Our goal is to produce and characterize mTLT-1 monoclonal antibodies from hybridoma cell lines to increase our understanding of TLT-1 in thrombotic and inflammatory diseases.
Materials and Methods
Cell culture maintenance and supernatant collection and processing
The immunization, hybridoma production, and initial screening were completed by the company Abcam. Rabbits were immunized with the peptides 94LQEEDTGEYGCMVEGA110 and 122PPVPGPREGEEAEDEK139 and tested positive for antibodies to TLT-1. Spleens were harvested; hybridomas made; and positive clones were identified by enzyme linked immunosorbent assay (ELISA). Nine clones were initially identified, and these clones could be grouped into two based on sequence identity (clones 4.6 and 4.8). Rabbit hybridoma clones 4.6 and 4.8 were maintained in a condition of semiadherence in RPMI-1640 Medium (Sigma), supplement A (Abcam), β-mercaptoethanol 0.05 mM (Calbiochem), and 10% fetal bovine serum (FBS; Sigma) at 37°C and 5% CO2. To increase antibody production, we gradually decreased the FBS percentage from 10% to 0%.(9) At high cell culture confluence (up to 90%), the cell culture medium was collected and centrifuged at 4°C, 400 g for 5 minutes to obtain the supernatant. We completed a second centrifugation at 4°C, 3000 g for 30 minutes, to remove any cells or cell debris that remained. Supernatants were stored at 4°C; long-term storage at −20°C.
Supernatant purification
Supernatant purification was performed with the Protein A agarose beads (Sigma) as the manufacturer protocol suggests. The aqueous suspension containing the beads was placed into Poly-Prep® Chromatography Column (Bio-Rad). To elute the antibody attached to the beads 500 μL of buffer B (0.2 M Na2HPO4, 0.1 M citric acid, and deionized H2O, at pH 2.7) was used. The eluates were neutralized using 5 M NaOH to pH 7.
Isolation of platelet-rich plasma and washed murine platelets
Peripheral blood was collected using cardiac puncture on anesthetized mice with a syringe containing 200 μL 3.8% sodium citrate. Anticoagulated blood was centrifuged at 900 rpm for 10 minutes at room temperature, and the upper 2/3 of platelet-rich plasma (PRP) was collected by aspiration. Prostaglandin E1 (0.5 μM) and apyrase (0.02 U/mL) were added to the PRP, and after each resuspension. Washed platelets were purified from PRP by centrifuge at 1350 g for 5 minutes. Platelet poor plasma was removed by aspiration, and the platelets were washed with 1 mL of Tyrode's buffer (134 mM NaCl, 2.9 mM KCl, 0.34 mM Na2HPO4, 1 mM MgCl2, 10 mM HEPES, 5 mM
Western blot
Washed platelets (3 × 108/mL) were lysed with lysis buffer (1% Triton-X 25 mM HEPES, 100 mM NaCl, 1 mM sodium orthovanadate, 10 ng/mL leupeptin, and 1 ng/mL aprotinin). Wild type (WT) and treml1−/− mouse platelet lysate aliquots were mixed with 2× loading dye (Bio-Rad), boiled at 94°C for 5 minutes, and ran on a sodium dodecyl sulfate (SDS)-polyacrylamide gel (4%–20% gradient Mini-PROTEAN® TGX Stain-Free™ Gels; Bio-Rad). After electrophoretic resolution, proteins were transferred to a polyvinylidene difluoride (PVDF) membrane (Bio-Rad) by Trans-Blot® Turbo™ Blotting System (Bio-Rad). The blotted membrane was then blocked for 1 hour at room temperature with Tris-buffered saline (TBS) containing 0.1% Tween-20 (TBST) and 5% (w/v) nonfat dry milk. The membrane was then incubated with supernatants directly (clone 4.6) or 1:1000 (clone 4.8) on a shaker overnight at 4°C. The membrane was washed and then incubated on a shaker at room temperature for 1 hour with donkey anti-rabbit horseradish peroxidase-conjugated (HRP) secondary antibody (Jackson ImmunoResearch). The secondary antibody was diluted 1:10,000 in 5% (w/v) nonfat dry milk TBST. Bands were visualized using Pierce™ ECL Western Blotting Substrate (Thermo Fisher Scientific) and by BioSpectrum® Imaging System (UVP, LLC).
Immunoprecipitation
Washed WT platelets were lysed with 1 mL of lysis buffer (1% Triton-X 25 mM HEPES, 100 mM NaCl, 1 mM sodium orthovanadate, 10 ng/mL leupeptin, and 1 ng/mL aprotinin). Supernatants (5 μL) from each clone were incubated with platelet lysate at 4
Flow cytometry
The platelets from WT and treml1−/−, prepared as above, at 3 × 107 platelets/μL were labeled with: 100 μL of hybridoma supernatant from each clone as primary antibody, or a polyclonal rabbit anti-mTLT-1(5,8) diluted 1:50 in FACS Buffer (0.1% BSA, 0.1% sodium azide, and phosphate-buffered saline (PBS) to 500 mL), which was used as positive control. The tubes were then incubated at 4°C for 20 minutes. Subsequently, platelets were washed with FACS buffer, and a goat anti-rabbit Fluorescein-5-isothiocyanate (FITC) secondary antibody and CD41-Phycoerythrin (PE) antibody (MWReg30; BD Biosciences®) were added to the tubes and incubated at 4°C for 20 minutes. After a wash step in FACS Buffer the pellet obtained was resuspended in 200 μL of FACS Buffer and counted by BD Accuri™ C6 cytometer (BD Biosciences). Whole blood flow cytometry was completed as in reference.(10)
Immunofluorescent staining
Round 12 mm coverslips (Warner Instruments) were coated with fibrinogen (Calbiochem®) at a concentration of 100 μg/mL for 1 hour at 37°C. Subsequently, 6 × 106 platelets were added per coverslip and allowed to spread for 30 minutes. Samples were washed with 1× PBS, permeabilized with BD Cytofix/Cytoperm™ (BD Biosciences) at 4°C for 20 minutes, and finally washed with 1× BD Perm/Wash™ (BD Biosciences) at 4°C for 20 minutes. The coverslips were incubated at room temperature for 1 hour with the primary antibodies. A rabbit anti-mTLT-1 antibody was diluted 1:200 in 1× BD Perm/Wash as positive control. Coverslips treated with the anti-mTLT-1 antibody produced by clone 4.8 and 4.6 were covered by a volume of 200 μL. After two washes in 1× PBS the coverslips were incubated at room temperature for 1 hour in the dark with goat anti-rabbit FITC conjugated (Jackson ImmunoResearch) and CD41-PE. The platelets were washed once in 1× PBS and were preserved with fluorescence mounting medium (Dako) and stored at 4°C in the dark. Pictures were taken with Nikon A1R Confocal Microscope at the Materials Characterization Center, Molecular Science Research Center (MSRC) of the University of Puerto Rico.
Aggregometry
The WT mouse platelet concentration used in this study was 2.0 × 108 cells/mL in a final volume of 400 μL, including 40 μL of hybridoma supernatants (clone 4.8 or 4.6) in the solution. Activation was achieved with 5 or 10 μM adenosine diphosphate (ADP) (Sigma). Platelet aggregation analysis was performed using a CHRONO-LOG® Model 700 Whole Blood/Optical Lumi-Aggregometer (Chrono-Log Corp).
Antibody sequencing
Approximately 5 × 106 hybridoma cells were lysed with 1 mL of TRIzol Reagent (Invitrogen) according to the manufacturer's protocol. The cDNA was prepared using the First Strand cDNA Kit (Invitrogen) according to the manufacturer's protocol. Primers were designed to target the conserved regions of rabbit IgG genes using heavy-chain primers (forward, 5′-ATGGAGACTGGGCTGCGCTGGC-3′; reverse, 5′-CTATTTACCCGGAGAGCGGGAG-3′) and light-chain primers (forward, 5′-ATGGACACGAGGGCCCCCACTC-3′; reverse, 5′-CTAACAGTCACCCCTATTGAAGC-3′). We used 200 μM of each primer; NovaTaq Polymerase master mix (Millipore Sigma) and polymerase chain reaction (PCR) conditions were as follows: Melting 95
Mice maintenance
Mice were housed at the University of Puerto Rico Animal House under the conditions of National Institutes of Health (NIH), and animal care was provided according to the procedures outlined in A Guide for the Care and Use of Laboratory Animals, 8th edition. Animals had regular diet and free access to water.
Mouse injury model
Procedures were approved by the IACUC at the University of Puerto Rico Rio Piedras. Eleven-week-old WT mice were used to assure an appropriate tissue response to skin injury.(12) The mice were anesthetized intraperitoneally with a solution with 100 mg/kg ketamine and 10 mg/kg xylazine. The dorsal areas to be burned on the left and right flanks were shaved to expose the skin. The burn assay was performed with a metallic tip warmed in a water bath to reach the temperature of 100°C. The metallic tip was pressed against the left and right dorsal flank for 5 seconds, to ensure proper injury of the skin exposed to the heat. For the first 3 days, 100 μL of the purified clone 4.8 was administered topically to the left side, and 100 μL of the neutralized elution solution was applied to the right side as negative control. The normalized wound area ratio was calculated for both sides, in each mouse. Results were graphed using GraphPad Prism (version 7.00 for Mac, GraphPad Software, La Jolla, CA,
Results
Light-chain sequence variation between clone 4.8 and clone 4.6 antibodies
Rabbits were immunized with two peptides whose sequences are derived from TLT-1. The first was “94LQEEDTGEYGCMVEGA110,” which is derived from the CDR loop 3. This peptide was chosen because the 17mer peptide encompassing these 16 amino acids blocks platelet function.(13) The second peptide was “122PPVPGPREG
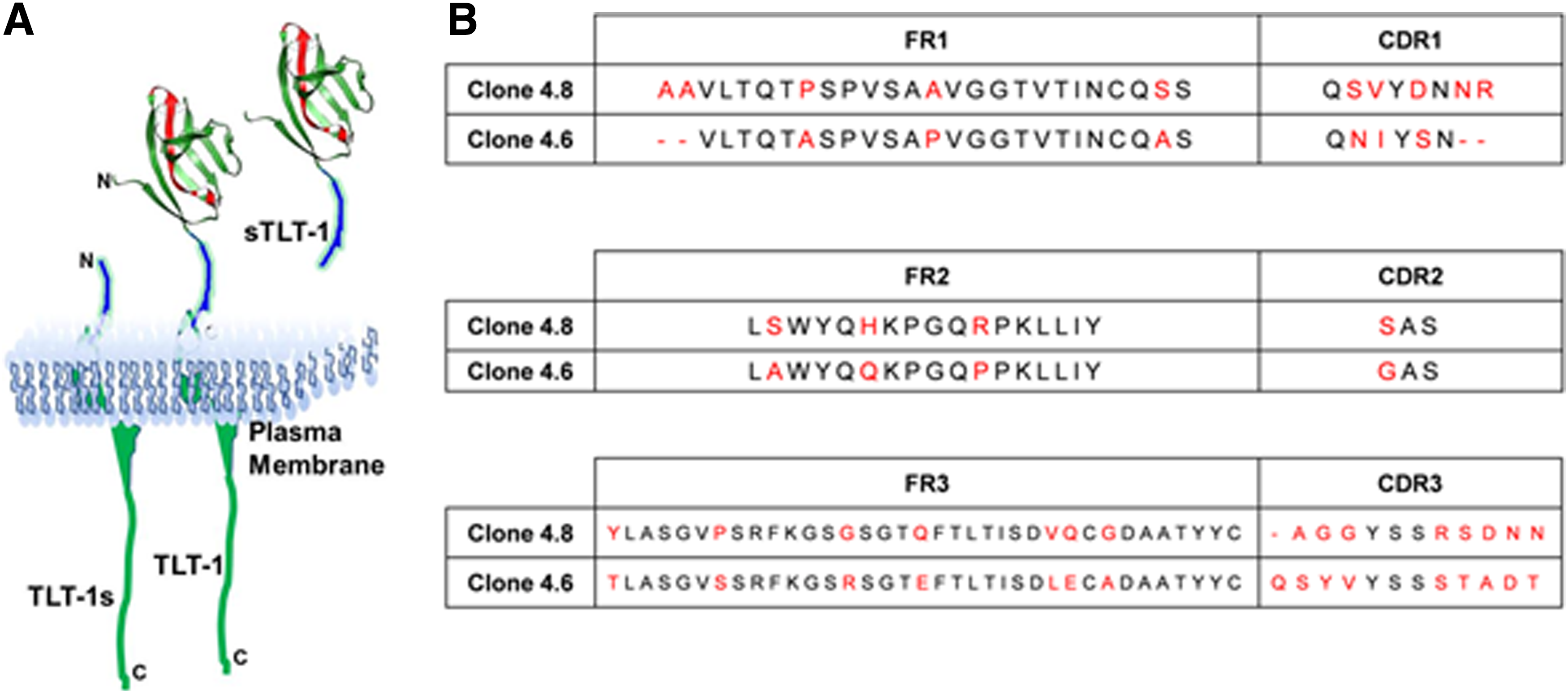
TLT-1 epitope and light chain alignment for clones 4.6 and 4.8.
Sequencing of the light and heavy chain antibody clones revealed that there were only two different clones denoted as clones 4.6 and 4.8. While there were significant differences in the light chains of these two clones, the heavy chains were identical (data not shown). The light chains of mTLT-1 antibody from both clones were amplified by PCR, and the amplicon for each antibody was sequenced and blasted against the IMGT germline V gene database with the software IgBLAST. Sequence similarity alignment (by COBALT)(11) of FR and CDRs is shown in Figure 1B for both clones. The clones in CDR loops 1 and 3 have striking differences between the two clones.
Antibody clone function for western blot and immunoprecipitation
First, we evaluated if these clones were functional in western blot analysis. The western blot analysis (Fig. 2A) on WT mouse platelet lysate showed a prominent band of ∼42 kDa, compatible with the molecular weight of mTLT-1 and confirmed by the control polyclonal anti-mTLT-1.(6) Both clones also recognized the soluble fragment (doublet at about 22 kDa)(5) and TLT-1s (∼27 kDa) shown for clone 4.8 in Figure 2B where human embryonic kidney 293 cells were either transfected with TLT-1s or mock transfected, and a band is seen only in the TLT-1s lane. As expected, no band was observed in the lanes of the treml1−/− mouse platelet lysate. Neither clone recognized human TLT-1.

Blot analysis using two antibody clones.
The immunoprecipitation (IP) assay (Fig. 2C) was performed on murine WT platelet lysates and mTLT-1 antibody from both clones. Immunoprecipitates were analyzed in nonreducing conditions. A thick band of ∼42 kDa is shown in the upper panel (Fig. 2C) for the sample treated with clone 4.8 and clone 4.6. The bands are consistent with the molecular weight of mTLT-1(6) and comparable to the whole cell lysate (WCL) positive control. IgG control showed only a faint band considered to be background. Afterward, the membrane was stripped, and an anti-rabbit HRP antibody was added to determine the relative amount of antibody in the supernatants. A thick band of ∼150 kDa is shown in the lower panel (Fig. 2C) for all IPs. As expected, no band is present in the WCL lane. These results indicate that the anti-mTLT-1 antibodies from both clones specifically bind to the mTLT-1 protein and are suitable for use in western blot and IP. The lower amount of TLT-1 that immunoprecipitated with clone 4.6 is consistent with the lower quantity of antibody in the supernatant as denoted by faint bands when blotted with the anti-rabbit HRP secondary antibody.
Antibody clones recognize TLT-1 on activated platelets by flow cytometry
The exposure of mTLT-1 on platelet membrane is a marker of platelet activation; therefore, we evaluated if we could use these clones to identify TLT-1 on the surface of activated platelets. Our results show, as expected, low baseline interaction of the antibodies with treml1−/− and resting wild-type platelets. A robust signal is generated upon incubation with either clone 4.6 or 4.8 on activated platelets (Fig. 3). Our results demonstrate that these clones specifically detect mTLT-1 on activated platelets and can be used to mark platelet activation. Fig. 3B shows that clone 4.8 works in whole blood flow cytometry assays. Clone 4.8 was subsequently purified and labeled with Alexa Fluor 647. Labeled clone 4.8 was incubated with whole blood for the denoted time points and measured by flow cytometry. Distinct levels of TLT-1 expression can be seen at each time point, and receptor shedding is evidenced with 5 μM/mL collagen and 10 nM thrombin by the decrease in MFI compared to 20 minutes.
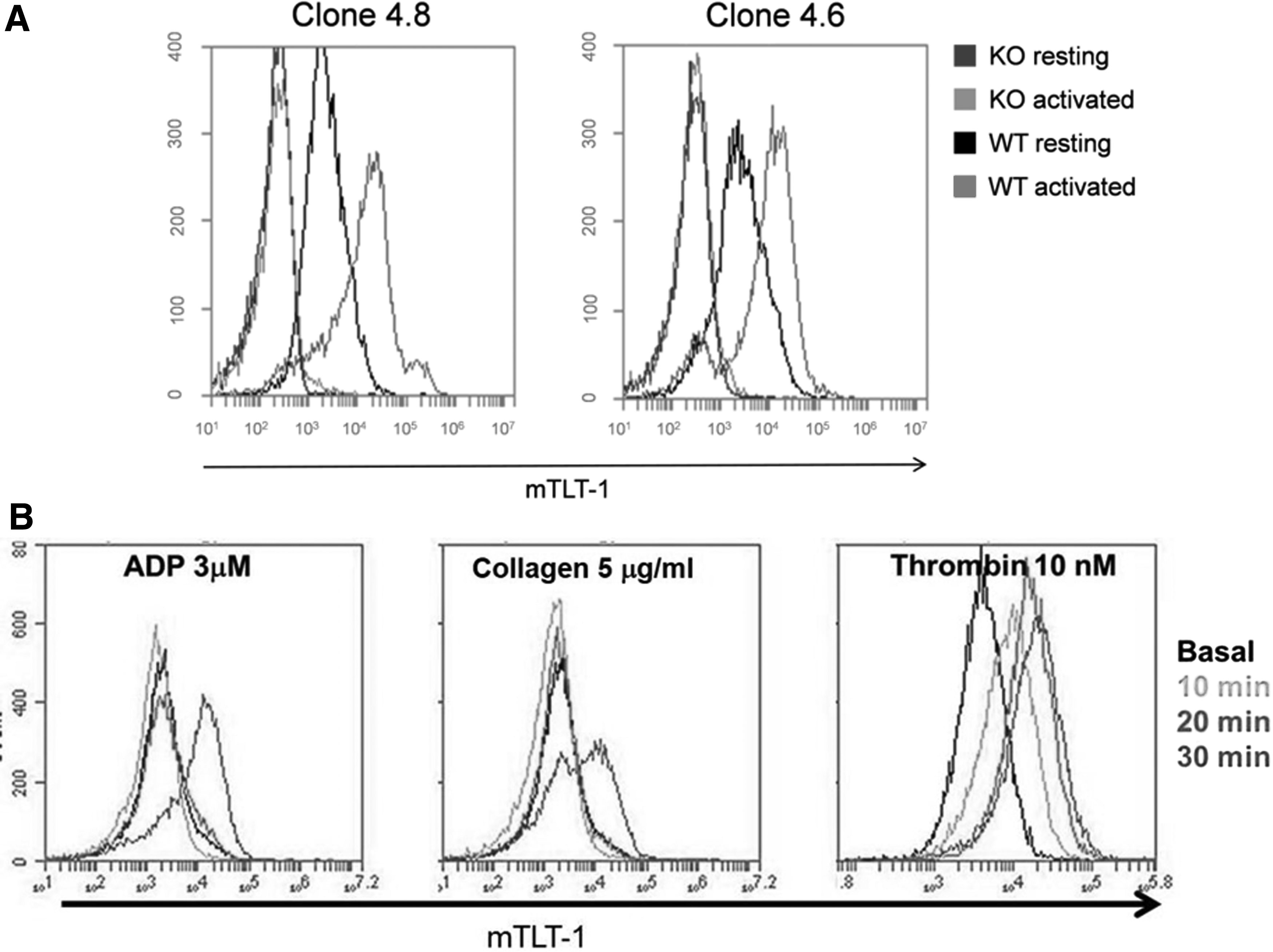
Clones 4.6 and 4.8 bind to activated WT platelets.
Evaluation of antibody clones for fluorescent microscopy
Next, we tested the ability of our antibodies to label platelets for immunofluorescence microscopy on WT and treml1−/− mouse platelets. Both clones 4.6 and 4.8 demonstrated specific labeling of resting and activated platelets from WT mouse (Fig. 4C, D). Some of the platelets showed punctate staining representative of TLT-1's storage in the α-granules, and others show TLT-1 dispersed throughout the cell. Our staining with these clones is consistent with the staining seen by the positive control (Fig. 4B). There was no appreciative staining seen with the treml1−/− platelet negative controls (Supplementary Fig. S1). Our data demonstrate that both clones can be used for immunofluorescence.
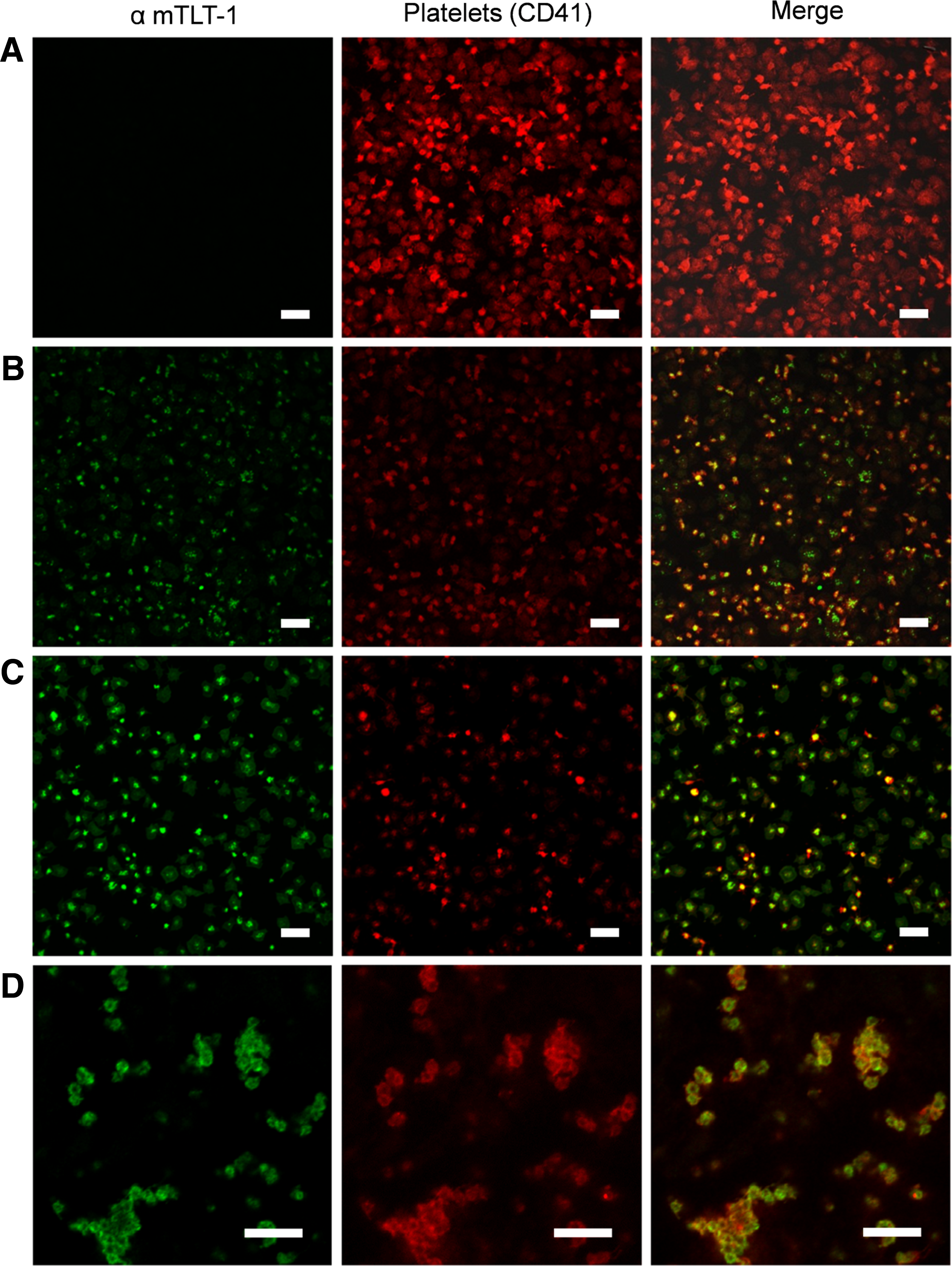
Fluorescent microscopy of murine platelets treated with hybridoma supernatants. Platelets were identified by an anti-mouse CD41-PE (red) and platelets stained with: secondary only as the negative control
Clones 4.8 and 4.6 inhibit platelet aggregation
Single-chain fragments to human TLT-1 have been shown to inhibit platelet aggregation. To investigate the effect of our antibodies on platelet aggregation, we performed an aggregometry assay. Clone 4.8 demonstrates inhibition with low dose thrombin, where clone 4.6 does not (Fig. 5A). At higher doses of thrombin, no inhibition was seen (data not shown). At lower ADP concentration both antibodies reduce platelet aggregation compared to the control (Fig. 5B). At higher ADP concentration the antibody from clone 4.6 is comparable to the control but the antibody from clone 4.8 is inducing platelet disaggregation (Fig. 5C). These results suggest that mTLT-1 antibodies from both clones reduce the platelet aggregation at low ADP concentration. At high ADP concentration mTLT-1 antibody from clone 4.8 appears to be affecting the second wave of reversible aggregation, typically associated with this agonist.(15)
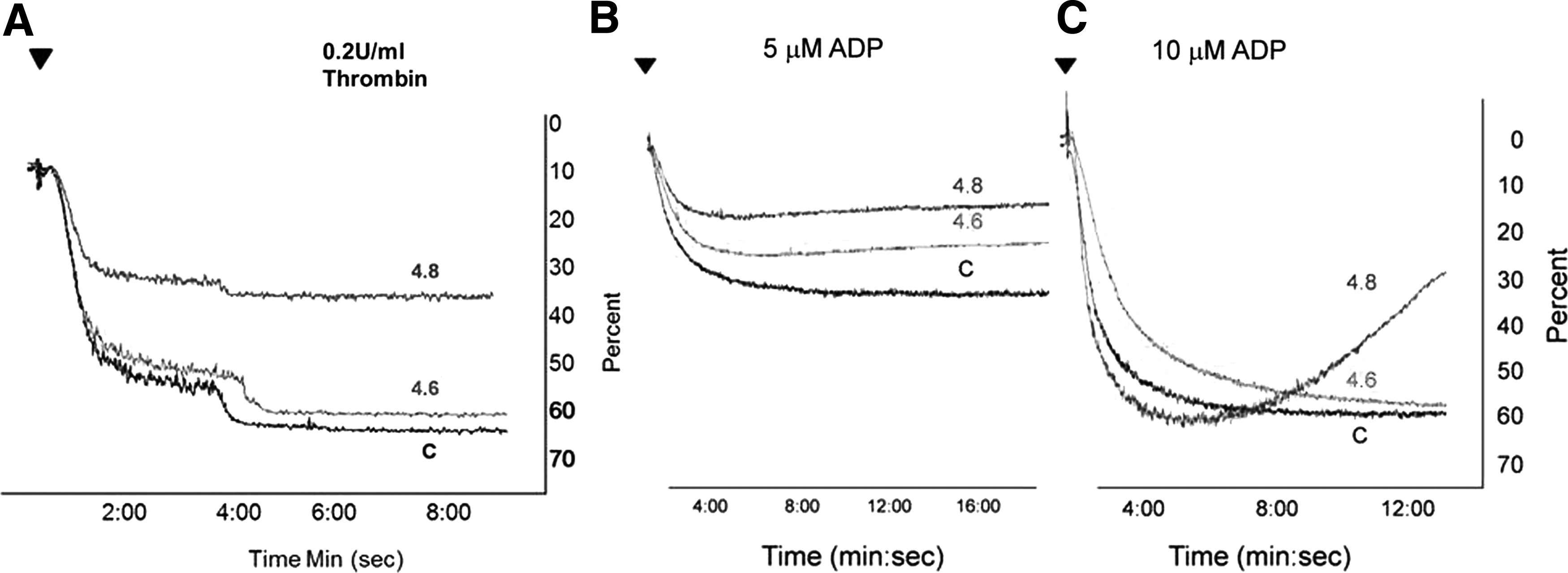
Antibody clones inhibit platelet activation. WT platelets were treated with purified supernatants, from both clones, and activated with 0.2 U/mL thrombin
Clone 4.8 antibody inhibits wound healing
Recent studies demonstrated that mTLT-1 positively affects inflammation and vascular hemostasis at the vascular injury site.(10) Based on our results described above and higher antibody productivity, we decided to test the effect of clone 4.8 antibody on wound healing. To this end, two thermal wounds on the murine dorsal area were produced. Our antibody was topically applied on the left side for the first 3 days; on the right side we applied the vehicle (Fig. 6A). Results show that by day 3, treated wounds were widely hyperemic, and the wound area was larger than the controls. Red granulation tissue was more evident by day 6 on wounds treated with mTLT-1 antibody. The formation of an epithelial layer and hair growth were absent on treated wounds; conversely, control wounds showed complete hair regrowth. Measurements of the wound show that on the third day of treatment with the antibody, the wounds were actually larger in area (Fig. 6B). Clearly, there are obvious visible differences between the wounds during the topical application period (day 1–3) and post-treatment period (day 4–6). Although the increase in wound size was not deemed significant, introduction of the antibody caused a delay in the completion of the healing of the wound (p = 0.02, n = 4).

mTLT-1 antibody from clone 4.8 affects wound healing process in WT mice.
Discussion
The characterization of TLT-1 receptor suggests that it plays a role in both hemostasis and inflammation, and further characterization of this receptor will lead to a better understanding of the process behind thrombus formation during hemostasis.(16) To develop a model that will allow us to investigate thrombus-related pathologies, we characterized two rabbit monoclonal antibodies. The hybridoma clones 4.6 and 4.8 produced the monoclonal antibodies specifically binding mTLT-1, as demonstrated by western blot and IP assay. The translocation of mTLT-1 to the platelet surface is a hallmark of platelet activation.(10) To evaluate the ability of these clones to recognize activated platelets, flow cytometry was performed on resting and activated platelets. Both antibodies positively bound to WT activated platelets showed limited binding to WT resting platelets. Both resting and activated treml1−/− platelets displayed low fluorescence intensity peaks, indicating an absence of interaction between our antibodies and mTLT-1, as expected. Clone 4.8 directly labeled with Alexa Fluor 647 worked well in whole blood flow cytometry with the ability to distinguish subtle changes in platelet activation and TLT-1 receptor shedding. Consistent with these results, we showed that these clones function in immunofluorescence as well. Taken together, these results demonstrate that we can identify mTLT-1 under multiple conditions with these clones, making them valuable molecular probes for our mouse model.
To assess the degree of functionality of our clones, we tested their effect on agonist-mediated platelet aggregation. The aggregometry results indicate that both clones inhibit platelet aggregation, but the mTLT-1 antibody from clone 4.8 appears to have a stronger effect. While this variance may be because of affinity differences between the two clones as denoted by the subtle changes in sequence, it is more likely due to lower concentration of clone 4.6 compared to 4.8 (data not shown). We were able to produce clone 4.8 more readily than clone 4.6. It is for this reason we pursued wound healing studies with 4.8 and not 4.6.
Previous studies suggested that TLT-1 is involved in inflammatory hemostasis,(10) but its effects on burn wound healing have not been determined. To investigate the role of mTLT-1 at the site of injury, we inflicted a thermal wound on murine skin. Topical application of clone 4.8 markedly retarded the wound healing process compared to controls. Specifically, during the third to sixth day the treated wound appeared to be more inflamed than the control. This is consistent with idea that platelets play an important role in the first 3 to 4 days of the wound. Approximated area measurements of the treated side supported an increased wound area and a delay in the wound healing compared to the control. These results suggest that mTLT-1 is necessary for a faster wound healing and proper scar formation.
In this study, we presented two mTLT-1 antibodies that can be efficiently used to identify platelet activation and to identify mTLT-1 during various assays in our mouse model. In addition, we demonstrated that our clone 4.8 antibody affects the wound healing during the regeneration process of scalded murine skin. Based on the inhibition of aggregation, we hypothesize that the antibody blocks TLT-1's interaction with fibrin(ogen) and inhibits platelet signaling that adds stability to the clot. Moreover, these results suggest that we may be able to use these clones to help identify TLT-1's mechanism of action with the goal of developing TLT-1 derived therapeutic treatments in inflammatory or hemostasis-based aliments.
Footnotes
Acknowledgments
The authors thank students Omar Rios and Franklin Staback for their suggestions on the western blot analysis and Elizabeth Evans for editing tips. A special thanks to Dr. Esther A. Peterson for sharing Trans-Blot Turbo (Bio-Rad). In addition, the authors thank Lic. Bismark Madera, from the Material Characterization Center, for the confocal microscope pictures and to the Molecular Science Research Center (MRSC, San Juan PR) and special thanks for the guidance given to them by the professors in Topicos 2017. Funding: Translational Study of Platelet Specific Receptor TREM-like transcript, TLT-1 Grant number: 1R01HL090933-01A2 Grant. RISE NIH Grant number: 5R25GM061151-16. LSAMP-Bridges to the Doctorate award number: 1612393, NIGMS-RISE R25GM061838 and U54MD007600.
Authors' Contributions
Cell maintenance and supernatant purification: B.M., J.M.-O., L.MD.-D., L.H.-M., and A.R.-D. Western blot: B.M. and J.M.-O. Immunoprecipitation: B.M., J.M.-O., L.M.D.-D., L.H.-M., and A.V.W. Flow cytometry: B.M., J.M.-O., L.M.D.-D., L.H.-M., and A.V.W. Immunofluorescence: B.M., J.M.-O., L.M.D.-D., L.H.-M., E.L.L.-D., M.K., and A.V.W. Aggregometry: B.M. and A.V.W. Sequencing: L.H.-M., J.M.-O., and A.V.W. Mouse injury model: B.M., L.M.D.-D., L.H.-M., D.B.-V., J.M.-P., and A.V.W. Statistics: E.S.-R. TLT-1/antibody modeling: K.C.C. and A.R.L. Article: B.M. and A.V.W. with the collaboration of J.M.-O., L.M.D.-D., L.H.-M., D.B.-V., J.M.-P., J.A.R.-C., and J.C.V.-S. Pictures: B.M., J.M.-O., L.M.D.-D., L.H.-M., J.A.R.-C., E.S.-R., and A.V.W. All authors read and approved the final article.
Author Disclosure Statement
A provisional patent covering all potential commercial and developmental uses of the antibodies described in this article has been obtained.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.