
Abstract
Malaria is a life-threatening infectious disease and continues to be a major public health crisis in many parts of the tropical world. Plasmodium falciparum is responsible for the majority of mortality and morbidity associated with malaria. During the intraerythrocytic cycle, P. falciparum releases three proteins with high histidine content as follows: histidine-rich protein 1 (HRP1), histidine-rich protein 2 (HRP2), and histidine-rich protein 3 (HRP3). Currently, most of the diagnostic tests of P. falciparum infection target HRP2, and a number of monoclonal antibodies (mAbs) against HRP2 have been developed for use in HRP2 detection and quantification. When parasites have HRP2 deletions, the detection of HRP3 could augment the sensitivity of the detection system. The combination of both HRP2 and HRP3 mAbs in the detection system will enhance the test sensitivity. In the HRP quantitative enzyme-linked immunosorbent assay (ELISA), both HRP2 and HRP3 contribute to the result, but the relative contribution of HRP2 and HRP3 was unable to investigate, because of the nonavailability of HRP3 specific antibody ELISA. Hence an ELISA test system based on HRP3 is also essential for detection and quantification. There is not much documented in the literature on HRP3 antigen and HRP3 specific mAbs and polyclonal antibodies (pAbs). In the present study, recombinant HRP3 was expressed in Escherichia coli and purified with Ni-NTA agarose column. The purified rHRP3 was used for the generation and characterization of monoclonal and pAbs. The purification of monoclonal and pAbs was done using a mixed-mode chromatography sorbent, phenylpropylamine HyperCel™. With the purified antibodies, a sandwich ELISA was developed. The sandwich ELISA method was explored to detect and quantify HRP3 of P. falciparum in the spent medium. The generated mAbs could be potentially used for the detection and quantification of P. falciparum HRP3.
Introduction
M
Therefore, it is crucial to understand the important parameters in the transmission of the disease and develop effective diagnostic strategies for its prevention and control. Today, rapid diagnostic tests (RDTs) are increasingly used for malaria diagnosis by detection of parasite biomarkers as they offer a result within 20 minutes. In these tests, plasmodium lactate dehydrogenase and fructose 1,6-bisphosphate aldolase (Aldolase) are commonly used as candidate targets for detection of infection with other Plasmodium species.(6–8)
During the intraerythrocytic cycle, P. falciparum releases three proteins with high histidine content—histidine-rich protein 1 (HRP1), histidine-rich protein 2 (HRP2), and histidine-rich protein 3 (HRP3).(9) HRP2 and HRP3 share similarities in their amino acid composition and sequence and share 85%–90% homology in nucleotide sequence.(10) These homologies have led to the proposal that genes for HRP2 and HRP3 have resulted from duplication and divergence from a common ancestral sequence and may complement each other in function.(11,12)
However, P. falciparum HRP2 and HRP3 is a biomarker that is predominantly used as a target for detection of P. falciparum infection.(9,11) HRP2 and HRP3 are the water soluble proteins that are produced by the asexual stages and gametocytes of P. falciparum. This protein is abundantly expressed in red cells, released during the rupture of infected red cells, and can remain in the blood for up to 28 days after the initiation of anti-malarial therapy, making it an excellent biomarker for diagnosis.(13–15) Sullivan et al.(16) have demonstrated heme polymerization in the presence of both HRP2 and HRP3.
In HRP2 detecting malaria RDTs, HRP3 also contributes to the detection of P. falciparum.(17) The polyclonal and monoclonal antibodies (mAbs) raised against HRP2 have been shown to cross-react with HRP3.(18,19) This is because HRP3 has amino acid sequence homology of more than 75% in the tandem repeat region to HRP2.(20) This cross-reactivity may not have a major impact on the sensitivity of the detection system.(21) But in situations of low-level parasitemia or when parasites have hrp2 gene deletions,(22) it could enhance the sensitivity of the detection system. Combination of HRP2 and HRP3 antibodies will enhance the test sensitivity.(20)
Therefore, new and more stable antigen binders represent important objectives in improving malaria RDT performance.
The aim of this study was to isolate new polyclonal antibodies (pAbs) and mAbs to PfHRP3 that may have better specificity, stability, and sensitivity than those currently used in RDTs. Polyclonal and mAbs against recombinant HRP3 were developed, purified, and characterized. A capture assay was developed for the detection and quantification of HRP3 of P. falciparum using purified polyclonal and mAbs.
Materials and Methods
Expression and purification of rHRP3
HRP3 gene was amplified from pRSETA-hrp3 and cloned in pET20b plasmid for rHRP3 expression. The plasmid (pET20b-hrp3) was transformed into Escherichia coli strain Rosetta 2 DE3 pLysS and was induced to express rHRP3 using 1 mM isopropyl-b-D-thiogalactopyranoside (IPTG). The recombinant proteins were harvested from the E. coli by freeze–thaw cycle and analyzed by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) to confirm the protein expression. Purification of rHRP3 was carried out by immobilized metal affinity chromatography using Ni-NTA agarose column (Qiagen, Hilden, Germany). The purified protein was analyzed for its purity by SDS-PAGE and was stored at −80°C until further use. As there were no specific antibodies available commercially for HRP3, we did an immunoblot analysis using commercially available HRP2 mAb (Span Diagnostics), as it is established to cross-react with HRP3 also.
Development of mAbs to rHRP3
Six-week-old female BALB/c mice were immunized subcutaneously with 100 mg of rHRP3 emulsified in Freund's complete adjuvant in the ratio (1:1). Booster injections were given thrice at an interval of three weeks, with 100 mg of rHRP3 emulsified in Freund's incomplete adjuvant. Three days before fusion, the mice were injected intraperitoneally with 300 mg of rHRP3 in saline. Spleen was excised from the mice; splenocytes were prepared as a single cell suspension and fused with Sp2/0 myeloma cells at a ratio of 5:1 using fusion reagent (50% w/v polyethylene glycol [PEG] in Dulbecco's modified Eagle's medium [DMEM] containing 10% dimethyl sulfoxide). Cells were then centrifuged, washed, and resuspended in DMEM supplemented with 20% fetal bovine serum, 50 mM b-mercaptoethanol, and HA (100 mM hypoxanthine, 5.8 mM azaserine). The fusion mixture was aliquoted in 96-well plates and placed in a 5% CO2 incubator at 37°C until the well contained visible growth of hybridoma cells. The screening of hybridomas secreting anti-rHRP3 mAbs was done by indirect enzyme-linked immunosorbent assay (ELISA). Selected hybridoma cell lines producing anti-rHRP3 mAbs were expanded and subsequently subcloned to monoclonality by the method of limiting dilution.
The isotypes of the mAbs produced by the selected hybridoma clones were determined using a Mouse mAb Isotyping Kit (Pierce, Rockford, IL) according to the manufacturer's protocols. The hybridoma cells were grown in a 175 cm2 cell culture flask containing DMEM with 10% fetal bovine serum. The cells were centrifuged at 400 g for 5 minutes, and the supernatant containing antibody was sterile filtered using 0.45 m filters (Millipore, Bellerica, MA).
Development of pAbs to rHRP3
Laboratory-bred female New Zealand albino rabbits were immunized with purified rHRP3, as described by Harlow and Lane.(23) The rabbits were immunized intramuscularly with 500 mg of purified rHRP3 protein emulsified in Freund's complete adjuvant, followed by booster doses of 500 mg of the antigen in Freund's incomplete adjuvant at an interval of 4 weeks. Animals were bled before immunization to be used as control. Serum samples were collected 10 days after the final booster and tested for immunoreactivity against the rHRP3 antigen by indirect ELISA.
Purification of mAbs and pAbs
Anti-rHRP3 mAbs from hybridoma culture supernatant and pAbs from rabbit antiserum were purified using phenylpropylamine HyperCel™ (Pall Life Sciences, Cergy, France).(24)
Quantification of HRP3 of P. falciparum using sandwich ELISA
Sandwich ELISA was standardized for rHRP3 with purified pAbs as capture antibody and mAbs as detection antibody to quantify HRP3 from P. falciparum spent medium. Microtiter plates were coated with capture antibody by adding 1 mg/100 mL/well of a 10 mg/mL solution in carbonate buffer (pH 9.6) and incubating overnight at room temperature. Plates were then blocked by adding 200 mL of phosphate-buffered saline (PBS) containing 5% skimmed milk and incubating the plates for 1 hour at 37°C. After three washes with wash buffer (PBS with 0.1% Tween-20 [PBST]), rHRP3 and P. falciparum spent medium (serially diluted) were added to the wells in four replicates, and the plates were incubated at 37°C for 2 hours. Detection was achieved with 10 mg/mL of mAbs, followed by goat antibodies to mouse IgG conjugated to peroxidase. Tetramethylbenzidine/H2O2 solution was used as a substrate for the enzyme with 2 M sulfuric acid as stopping solution. Absorbance was read at 450 nm with the multiwall plate reader. The results were expressed as mean absorbance at 450 nm for each group–standard deviation. The HRP3 concentration was determined by interpolating the ELISA absorbance of serially diluted P. falciparum spent medium against a stock of rHRP3 protein with known protein concentration. The HRP3 antigen level in sandwich ELISA assays was interpolated from a four parameter standard curve using the four parameter logistic curve online data analysis tool MyAssays Ltd.
Protein quantification
The total protein concentration of the antigen and antibody samples was quantified by Bradford's method(25) using bovine serum albumin (BSA) as reference protein for antigen work and IgG as reference protein for antibody work.
Result
Recombinant HRP3 was expressed using E. coli Rosetta 2 DE3 pLysS. Exploring the very high amount of histidine residues present in the protein, it was purified using Ni-NTA agarose column. mAbs against rHRP3 were obtained by the fusion of Sp2/0 myeloma cells and the spleen cells from rHRP3 immunized BALB/c mice. From the fusion experiments, wells that showed confluence growth of hybridomas were initially screened by means of indirect ELISA for the secretion of antibodies to rHRP3. The positive hybridomas were subcloned to monoclonality by means of limiting dilution.
The pAb against rHRP3 was developed in female New Zealand rabbits. The purified mAbs and pAbs were used for sandwich ELISA to quantify P. falciparum HRP3. Sandwich ELISA was standardized for rHRP3 to quantify native HRP3 from P. falciparum spent medium.
Sandwich ELISA using polyclonal as a capture antibody and monoclonal as a detection antibody showed linear range of absorbance (450 nm ± standard deviation) from 0.234 ± 0.016 to 1.465 ± 0.024 corresponding to 0.244 to 1000 ng/mL of rHRP3 (Table 1 and Fig. 1). The coefficient of variation at each target concentration ranged from 0.224% to 8.5%. Similarly, HRP3 from P. falciparum spent medium (double dilution 1: 8192) was quantified as 2421 ng/mL.
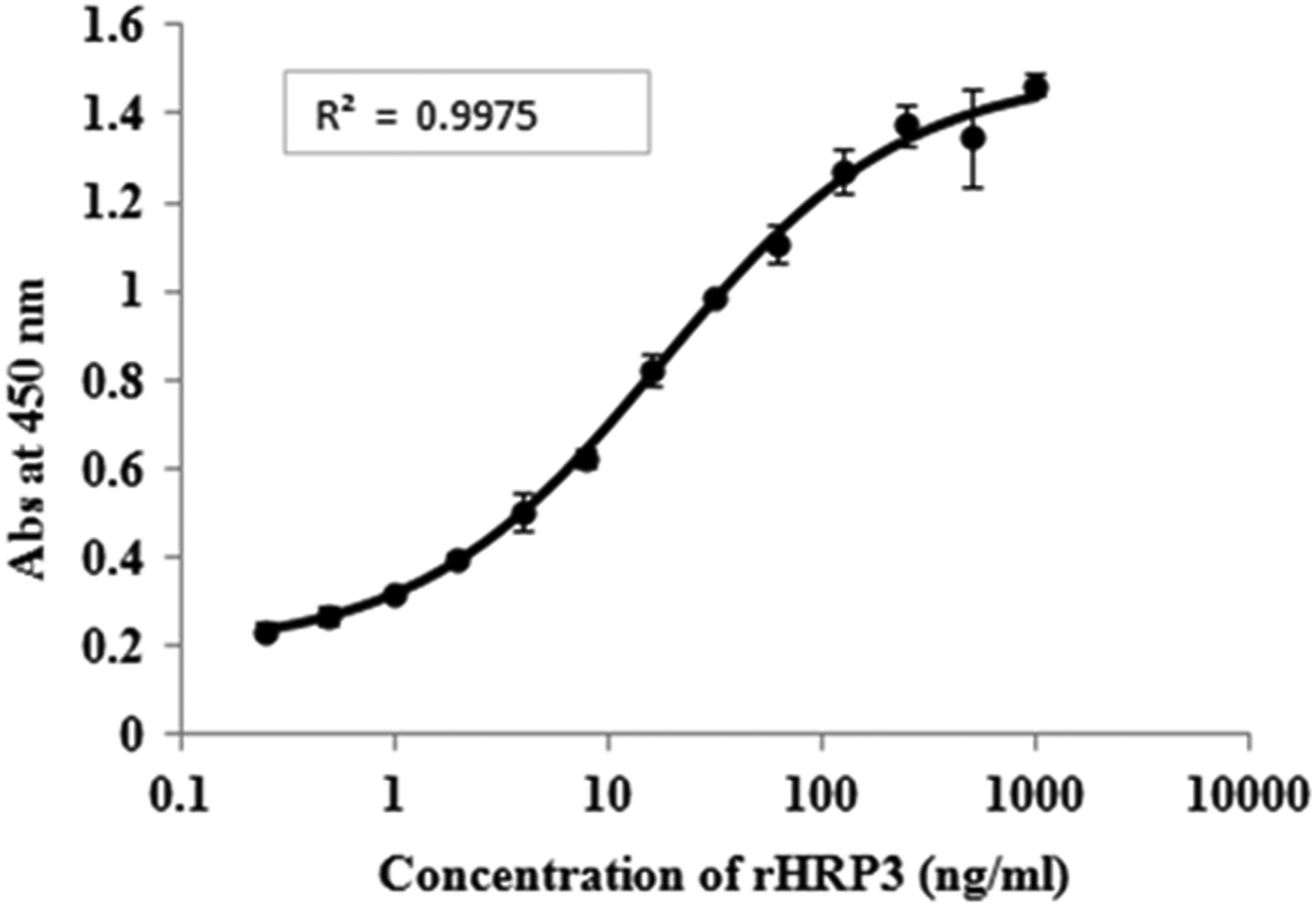
Four parameter logistic curve fit for sandwich enzyme-linked immunosorbent assay using polyclonal as capture antibody and monoclonal as detection antibody for rHRP3.
OD, optical density; SD, standard deviation; CV, coefficient of variation.
Discussion
Most of the diagnostic tests of P. falciparum infection target HRP2, and for detection and quantification of HRP2 a number of mAbs have been developed.(22)
The mAbs raised against HRP2 have been shown to cross-react with HRP3.(20,21) This is because HRP3 has an amino acid sequence homology of more than 75% in the tandem repeat region to HRP2.(22) Thus HRP3 also contributes to the detection of P. falciparum infections in HRP2 detecting malaria RDTs.(19) Because of the low abundance of HRP3 this cross-reactivity may not have a major impact on the sensitivity of the detection system.(23) But in situations of low-level parasitemia or when parasites have hrp2 gene deletions,(24) it could enhance the sensitivity of the detection system. Combination of HRP2 and HRP3 mAbs will enhance the test sensitivity.(22) In the HRP quantitative ELISA both HRP2 and HRP3 contribute to the result, but it is not possible to investigate the relative contribution of HRP2 and HRP3 because of the nonavailability of HRP3 specific antibody ELISA.(23) In this connection the HRP3 based ELISA quantification system is needed.
In the present study, monoclonal and pAbs against recombinant HRP3 were developed and used in the capture assay for the detection and quantification of HRP3 of P. falciparum.
Footnotes
Author Disclosure Statement
No competing financial interests exist.