
Abstract
T cell immunoglobulin and ITIM domains (TIGIT) is an inhibitory immunoreceptor expressed on NK cells, effector and memory T cells, and regulatory T cells (Tregs). The ligands for TIGIT are CD155 (PVR) and CD112 (PVRL2, nectin-2), which are broadly expressed on hematopoietic cells and nonhematopoietic cells. TIGIT negatively regulates antitumor responses, but promotes autoimmune reaction. Although neutralizing anti-human TIGIT mAbs are under clinical trials for cancers, how the blockade of TIGIT interaction with the ligands shows tumor immunity still remains unclear. Although analyses of mouse tumor model using a neutralizing anti-mouse TIGIT (mTIGIT) mAbs should be useful to address this issue, there are limitations to this type of studies due to unavailability of neutralizing anti-mTIGIT mAbs. In this study, we generated five clones of anti-mTIGIT mAbs, designated TX99, TX100, TX103, TX104, and TX105. We show that TX99 and TX100 showed the strongest binding to TIGIT. We also show that TX99 interfered with the interaction between TIGIT and CD155 and increased NK cell-mediated cytotoxicity against CD155-expressing RMA-S cells. Thus, TX99 is a unique neutralizing mAb that can be used for studies of mTIGIT functions.
Introduction
T
TIGIT negatively regulates antitumor response. Blockade of TIGIT interaction with the ligands enhances cytotoxicity against tumors by CD8+ T cells and NK cells.(3,10,11) On the other hand, although TIGIT-deficient mice do not develop spontaneous autoimmunity, they are highly susceptible to induced autoimmune diseases, such as experimental autoimmune encephalitis in mice.(12,13) Although anti-human TIGIT mAbs are under clinical trials for cancers, how the blockade of TIGIT interaction with the ligands shows tumor immunity still remains unclear. Although analyses of mouse tumor model using a neutralizing anti-mTIGIT mAb should be useful to address this issue, there are limitations to this type of studies due to unavailability of anti-mTIGIT mAbs.
In this study, we generated five clones of anti-mTIGIT mAb, named TX99, TX100, TX103, TX104, and TX105. In the present study, we examined the functional characteristics of TX99, which showed the strongest binding to TIGIT.
Materials and Methods
Animals
Wild-type mice (C57BL/6) and rats (Wister) were purchased from CLEA Japan, Inc. (Tokyo, Japan). Foxp3eGFP mice were provided by B. Malissen (UM2 Aix-Marseille Université).(14) All experiments were performed according to the guidelines of the Animal Ethics Committee of the Laboratory Animal Resource Center, University of Tsukuba.
Establishment of transfectant and fusion protein
Mouse Tigit cDNA tagged with the flag-encoding sequence at the 5′-end was subcloned into the retrovirus vector pMX. BW5147 and Ba/F3 cells stably expressing Flag-tagged mTIGIT were generated, as described.(15) The chimeric proteins consisting of the extracellular portion of mTIGIT or mouse CD155 (mCD155) fused with human IgG Fc portion at the C-terminus (TIGIT-Fc or CD155-Fc, respectively) were generated, as described.(6)
Establishment of hybridoma
On day 0, BW5147 transfectant expressing mTIGIT were emulsified with complete Freund's adjuvant (CFA) and the mixture was injected into the footpad of a rat for immunization. On day 29 and 36, mTIGIT-Fc fusion protein was emulsified with CFA and the mixture was injected into the footpad and the tail base for challenge. The popliteal, inguinal, and internal iliac lymph nodes were harvested and fused with SP2/O myeloma cells. The fused cells were cultured in HAT medium, and colonies were picked up and expanded in Medium E (Stem Cell Technologies, Vancouver, Canada). mTIGIT-expressing Ba/F3 transfectant or parental Ba/F3 were stained with culture supernatants of clones that were picked up and analyzed by flow cytometry for screening for anti-TIGIT mAb. Five clones, named as TX99, TX100, TX103, TX104, and TX105, which specifically recognized mTIGIT, were selected.
Flow cytometry analysis
Ba/F3 transfectant expressing mTIGIT or Ba/F3 cells were stained with TX99, TX100, TX103, TX104, and TX105 or isotype-matched control antibody, followed by secondary antibodies. For primary T cells, splenic mononuclear cells derived from Foxp3eGFP mice were stained with anti-NK1.1 (PK136), anti-CD3 (145-2C11), anti-CD8 (53-6.7) (BioLegend, San Diego, CA), and anti-CD4 (RM4-5) mAbs (BD Biosciences, San Jose, CA). CD4+GFP+ cells were identified as Treg. For activation of T cells, MACS (Miltenyi Biotec, Bergisch Gladbach, Germany)-sorted CD4+ or CD8+ cells were stimulated with anti-CD3 mAb (1 μg/mL) (145-2C11; TONBO Biosciences, San Diego, CA) and anti-CD28 (1 μg/mL) mAb (37.51; BD Biosciences) for 3 days. For activation of NK cells, mice were treated with poly I:C for 18 hours. Primary T cells and NK cells were stained with biotinylated mAbs, followed by fluorescein-conjugated streptavidin. Flow cytometry analysis was performed by using LSRFortessa flow cytometers (BD Biosciences). FlowJo software (Tree Star, OR) was used for data analysis.
Competitive binding assay
mTIGIT-expressing Ba/F3 transfectants were pretreated with the saturation dose of phycoerythrin (PE)-conjugated 1G9 (TONBO Biosciences), then stained with biotinylated TX99, followed by allophycocyanin (APC)-conjugated streptavidin (BD Biosciences), and analyzed by flow cytometry. Conversely, mTIGIT-expressing Ba/F3 transfectants were pretreated with the saturation dose of biotinylated TX99, then stained with PE-conjugated 1G9, and analyzed by flow cytometry.
Blocking assay
Ba/F3 transfectants expressing mTIGIT were incubated with mCD155-Fc fusion protein for 30 minutes at 4°C, washed with phosphate-buffered saline, and treated with purified TX99, 1G9 (BioLegend) or isotype-matched control Ab for 30 minutes at RT, followed by PE-conjugated anti-human IgG Fc secondary mAb (BioLegend). Cells were analyzed by flow cytometry.
Reporter assay
NFAT-GFP reporter cells were kindly supplied by Dr. Hisashi Arase (Osaka University).(16) Extracellular region of mouse Tigit and intracellular region of Fcer1g chimric constract were subcloned into the retrovirus vector pMX and transfected into the NFAT-GFP reporter cells. The reporter cells expressing mTIGIT-FcRγ were stimulated with plate-bound (10 μg/mL) or soluble-form TX99 or isotype-matched control Ab. GFP expression was analyzed by flow cytometry 24 hours after stimulation.
Cytotoxicity assay
C57BL/6 mice were treated with 250 ng Poly I:C. Eighteen hours later, the spleen was excised from the mice and the spleen cells were treated with anti-mouse CD16/CD32 Ab (2.4G2; TONBO Biosciences) at RT for 10 minutes, and then treated with TX99 or control Ab (0.01 − 10 μg per 1 × 106 cells) at 4°C for 30 minutes. These splenic mononuclear cells (1 × 106) were cocultured with RMA-S transfectant expressing mCD155 (1 × 105) in the presence of APC-conjugated anti-mouse CD107a mAb (BioLegend) and BD GolgiStop™ protein transport inhibitor (BD Biosciences) for 5 hours at 37°C in a 5% CO2 incubator. After coculture, these cells were stained by fluorescein-conjugated anti-mouse-NK1.1 and PE-Cy7-conjugated anti-mouse-TCRβ mAbs (BioLegend), followed by intracellular staining with PE-conjugated anti-mouse IFN-γ mAb (BD Biosciences) according to the manufacturer's instructions (Fix & Perm; Thermo Fisher Scientific, Waltham, MA) and then cells were analyzed by flow cytometry.
Results and Discussion
We generated five clones of anti-mTIGIT mAbs, designated TX99 (rat IgG1), TX100 (rat IgG1), TX103 (rat IgG2a), TX104 (rat IgG2a), and TX105 (rat IgG2a). These mAbs specifically bound to Ba/F3 transfectant expressing mTIGIT, but not parental Ba/F3 cells (Fig. 1A), indicating that the mAbs are specific to mTIGIT. Next, we examined the reactivity of these mAbs to spleen cells. Consistent with the previous report,(1–3) these mAbs specifically stained activated CD4+ T cells and CD8+ T cells, but not naive CD4+ T cells and CD8+ T cells (Fig. 1B, C). Tregs and NK cells were stained with these mAbs even in steady state and more strongly in activated state (Fig. 1B, C).
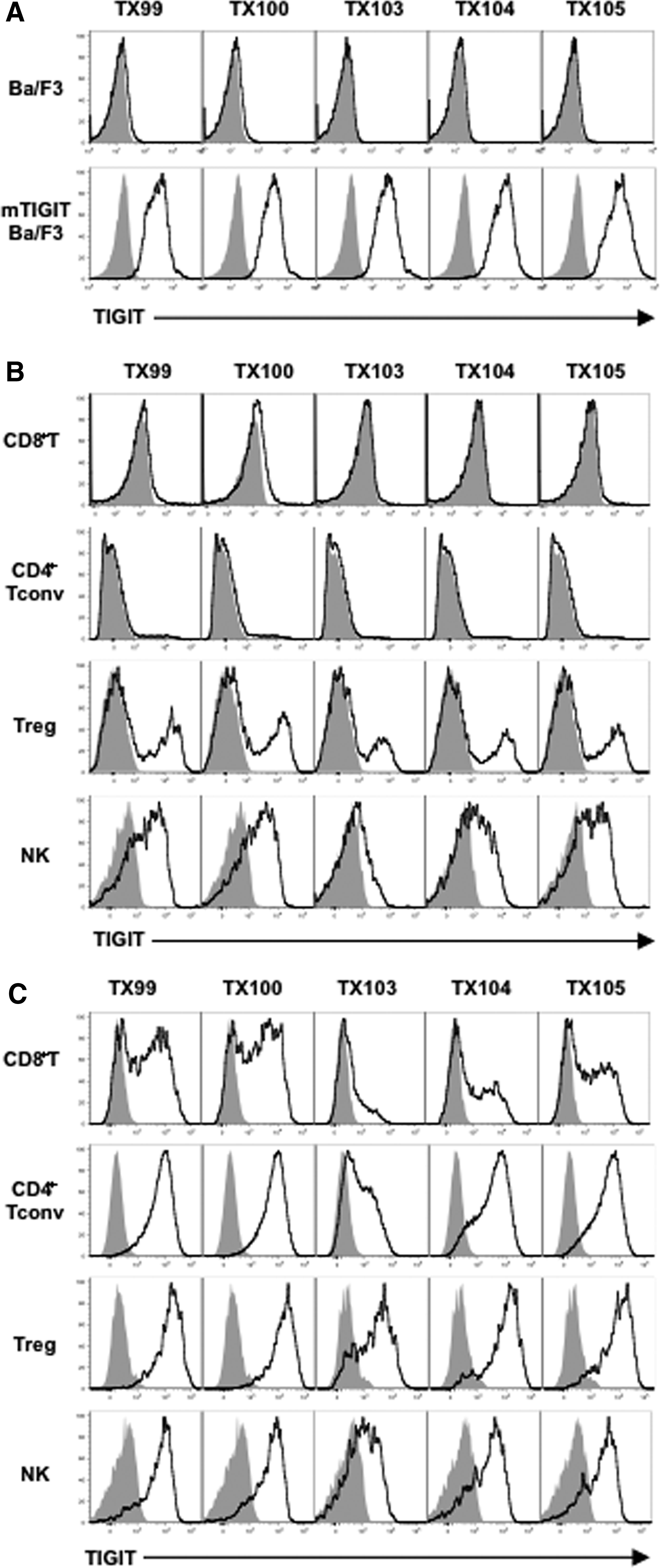
TX99 specifically recognizes mTIGIT.
Since TX99 and TX100 most strongly bound to TIGIT on spleen cells, we focused on TX99 for further analyses. To examine whether TX99 is able to induce TIGIT-mediated signal, we used NFAT-GFP reporter cells expressing TIGIT, in which GFP expression is induced by signaling through TIGIT (Fig. 2A). The reporter cells were stimulated with a soluble form of TX99. However, we did not detect any GFP expression, while GFP expression was induced when TIGIT on the reporter cells were crosslinked with plate-bound TX99 (Fig. 2B), indicating that TX99 does not have an agonistic activity.

Functional characteristics of TX99.
We next compared the functional characteristics of TX99 with 1G9, which is currently an only commercially available neutralizing anti-mTIGIT mAb.(17) Ba/F3 transfectants expressing TIGIT were pretreated or not with a saturated dose of 1G9 and then treated with biotinylated TX99, followed by APC-conjugated streptavidin. Whereas TX99 significantly inhibited the binding of 1G9 to Ba/F3 transfectants expressing TIGIT, the effect of 1G9 pretreatment on the binding of TX99 was scarcely observed (Fig. 2C). These results suggest that the affinity of TX99 for TIGIT seems to be higher than that of 1G9.
To examine whether TX99 and 1G9 interfere with the interaction between TIGIT and CD155, Ba/F3 transfectants expressing TIGIT were pretreated with mCD155-Fc fusion protein at a saturated dose, and then treated with TX99, 1G9, or control antibody, followed by PE-conjugated anti-human IgG mAb. Both TX99 and 1G9 inhibited the binding of CD155-Fc fusion protein to Ba/F3 transfectants expressing TIGIT in a dose-dependent manner (Fig. 2D), suggesting that TX99 as well as 1G9 interfere with the interaction between TIGIT and CD155. Of note, when we compared TX99 with 1G9, the inhibition of the binding of CD155-Fc fusion protein to Ba/F3 transfectants expressing TIGIT by TX99 was larger than that by 1G9 (Fig. 2D). TX99 was also able to upregulate NK cell-mediated cytotoxicity, which was evaluated by CD107a expression and IFN-γ production, against RMA-S transfectant expressing CD155 (Fig. 2E), consistent with previous reports.(2,3) Altogether, our studies demonstrated that TX99 is a unique neutralizing mAb that efficiently interferes with the interaction between TIGIT and CD155.
Footnotes
Acknowledgments
The authors thank S. Tochihara and W. Saito for their secretarial assistances and T. Nabekura and G. Okumura for discussion. This study was supported in part by grants provided by the Ministry of Education, Culture, Sports, Science, and Technology of Japan (Grant number 16H06387 to A.S. and Grant number 16H05169 to K.S.).
Author Disclosure Statement
No competing financial interests exist.