
Abstract
Staphylococcus aureus secretes a family of exoproteins structurally homologous to bacterial superantigens, such as toxic shock syndrome toxin-1 (TSST-1), and those exoproteins are thus called staphylococcal superantigen-like proteins (SSLs). Recent studies have revealed that SSLs play roles in evasion of the host defense by disturbing host immune responses. We previously reported that staphylococcal superantigen-like protein 5 (SSL5; a member of the SSL family) inhibited matrix metalloproteinase-9 (MMP-9), which is crucial for leukocyte recruitment to sites of infection. In this study, we established a mouse hybridoma clone (30G5C) producing a monoclonal antibody specific for SSL5. In immunoblotting analysis using recombinant His-tagged SSL1 to SSL14 (His-SSLs), the antibody was found to specifically recognize SSL5 without crossreactivity with other His-SSLs. The antibody bound to the C-terminal region of SSL5 (β-grasp domain), but did not interfere with the binding of SSL5 to MMP-9, suggesting that this antibody is useful for identification of SSL5-producing S. aureus and screening for inhibitors of the SSL5/MMP-9 complex formation.
Introduction
S
Recent studies have revealed that SSLs act as immune modulators to escape from the host immune defense; for example, SSL3 as a Toll-like receptor 2 (TLR2) antagonist,(6,7) SSL7 as an inhibitor for IgA-mediated opsonization and complement activation,(8) and SSL10 as an inhibitor for chemokine and complement functions.(9–11) Previous studies, including ours, have indicated that staphylococcal superantigen-like protein 5 (SSL5) binds to sialic acid-containing carbohydrate(s) of P-selectin glycoprotein ligand-1 (PSGL-1) and matrix metalloproteinase-9 (MMP-9), causing impairment of leukocyte migration to inflammatory tissues and hemorrhagic sites.(12–14) In addition, SSL5 binds to and activates platelets through interaction with cell membrane glycoproteins (GPIb and GPVI), which causes symptoms related to disseminated intravascular coagulation (DIC).(15,16)
Each SSL member possesses a common domain structure shared with TSST-1 and SPEC(17,18); an N-terminal β-barrel globular domain (oligonucleotide/oligosaccharide-binding fold, OB-fold) and a C-terminal β-grasp motif. The amino acid sequence identities of SSL1-SSL14 with SSL5 [NCBI-ProteinID: YP_498976] range from 23% to 48% (42% for SSL1 [YP_498971], 39% for SSL2 [YP_498972], 36% for SSL3 [YP_498973], 38% for SSL4 [YP_498975], 40% for SSL6 [YP_498978], 40% for SSL7 [YP_498979], 39% for SSL8 [YP_498980], 37% for SSL9 [YP_498981], 36% for SSL10 [YP_498982], 48% for SSL11 [YP_498986], 23% for SSL12 [YP_499668], 24% for SSL13 [YP_499669], and 24% for SSL14 [YP_499670]). SSL5 contains a carbohydrate-binding motif, which is highly conserved among SSL2, SSL3, SSL4, SSL6, and SSL11.(19) Therefore, a monoclonal antibody specific for SSL5 could provide a powerful tool to discriminate SSL5 from other SSLs. In this study, we established a hybridoma clone producing an anti-SSL5 monoclonal antibody and characterized its binding ability and potential applications.
Materials and Methods
Reagents
Restriction endonucleases were purchased from (TaKaRa Bio, Inc., Shiga, Japan). Isopropyl-β-
Recombinant SSL proteins
The genes for SSLs (SSL1 to 14) were amplified by PCR using primeSTAR MAX DNA polymerase (TaKaRa Bio) and genomic DNA of Staphylococcus aureus (ATCC 27733) as a template. The amplified DNA for full-length SSL5 (amino acid residues 32–234) was inserted into a pGEX-5X-1 vector (GE Healthcare) as described previously.(14) The truncated forms of SSL5, SSL5Nterm (amino acid residues 32–137, including the OB-fold domain), and SSL5Cterm (amino acid residues 128–234, including the β-grasp domain), were also prepared by inserting PCR-amplified SSL5Nterm and SSL5Cterm fragments into the BamHI/SalI and BamHI/NotI sites of a pGEX-5X-1 vector, respectively. The primers used for PCR were as follows: 5′-GGG GAT CCC CGA ACA TAA AGC AAA ATA T-3′ and 5′-GGG GTC GAC GTT TAT ATA ATC ATA ATA AGC-3′ for SSL5Nterm, and 5′-GGG ATC CCC AAT CAA GAC GCT TAT TAT GAT TAT A-3′, and 5′-GGG CGG CCG CTT ATC TAA TGT TGG CTT C-3′ for SSL5Cterm. Recombinant glutathione S-transferase (GST)-fused proteins were expressed in Escherichia coli (SoluBL21; Genlantis, San Diego, CA) in the presence of IPTG (0.2 mM at a final concentration) for 16 hours at 25°C, and purified by glutathione agarose according to the manufacturer's instructions. His-tagged SSLs (SSL1 to SSL14) were expressed in E. coli as described previously.(6) The DNA sequences were determined by the dideoxy method (Fasmac DNA Sequence Service, Tokyo, Japan).
Mice
BALB/c mice at 6 weeks of age were purchased from Japan SLC, Inc. (Shizuoka, Japan). All procedures for the experiments using mice were conducted in accordance with the Guide for the Care and Use of Laboratory Animals of the Hoshi University School of Pharmacy and Pharmaceutical Sciences.
Establishment of hybridoma
Monoclonal antibodies specific for SSL5 or GST were generated by fusing PAI myeloma cells (Japanese Collection of Research Bioresources, Ibaraki, Japan) with splenocytes of BALB/c mice immunized with a recombinant GST-SSL5. The immunizations of mice were performed as described previously.(20) Hybridomas were selected in hypoxanthine/aminopterin/thymidine (HAT) medium (Sigma-Aldrich), and the culture supernatants were screened by enzyme-linked immunosorbent assay (ELISA) using GST-SSL5 and GST. Monoclonal antibodies from clone 30G5C (IgG1/κ) against SSL5 and from clone 28A7 (IgG2a/κ) against GST were selected in this study. These hybridomas were cultured in serum-free medium (ASF medium 104; Ajinomoto Co., Inc., Tokyo, Japan), and antibodies were purified with a HiTrap Protein G HP column (GE Healthcare). The isotypes of monoclonal antibodies were determined by using the IsoStrip Mouse Monoclonal Antibody Isotyping Kit (Roche Diagnostics, Indianapolis, IN).
Purification of MMP-9
MMP-9 was purified from conditioned medium of PMA-stimulated THP-1 cells (a human monocytic leukemia cell line) essentially as described previously.(14,21) THP-1 cells were cultured with 5 nM PMA in ASF 104 serum-free medium for 2 days. The conditioned medium was mixed with gelatin Sepharose 4B and incubated for 4 hours at 4°C with gentle agitation. The beads were washed three times with TNC buffer (50 mM Tris-HCl, 100 mM NaCl, 10 mM CaCl2, pH 7.5). MMP-9 bound to the beads was eluted with TNC buffer containing 5% DMSO and immediately applied to a PD-10 column (GE Healthcare) to remove DMSO.
Immunoblotting and immunoprecipitation
Sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and immunoblotting were conducted as described previously.(22) Anti-SSL5 (30G5C) or anti-GST (28A7) antibody (1 μg) was incubated with Dynabeads Protein G (10 μL) with gentle agitation in a rotator for 30 minutes at room temperature. The Dynabeads Protein G/antibody conjugate thus obtained was mixed with purified GST-SSL5 or GST for 1 hour, and the mixture was incubated at room temperature for 1 hour. The beads were washed three times with phosphate-buffered saline (PBS) containing 0.5% Brij 35 to remove unbound proteins. Proteins bound to the beads were recovered by the treatment with SDS sample buffer (50 mM Tris-HCl, pH 6.8, 1% SDS, 5% glycerol, 0.01% bromophenol blue, 5% 2-mercaptoethanol) at 95°C for 3 minutes and subjected to SDS-PAGE.
Pull-down assay
Purified recombinant GST-SSL5 (1 μg) was bound to glutathione agarose (15 μL) at room temperature for 1 hour. The GST-SSL5-conjugated glutathione agarose was incubated with or without antibodies (1 μg; anti-SSL (IgG1/κ), anti-α3A integrin (IgG1/κ, isotype-matched control),(23) or anti-GST (IgG2a/κ) antibodies) at room temperature for 1 hour, and then with purified MMP-9 (1 μg) at room temperature for 2 hours. After the beads were washed with PBS containing 0.5% Brij 35, proteins bound to the beads were recovered with SDS sample buffer.
Results and Discussion
Establishment of hybridomas producing anti-SSL5 and anti-GST antibodies
First, we purified the recombinant GST-fused SSL5 from E. coli to be used as an immunogen. Purified GST-SSL5 gave a single band at ∼48 kDa in SDS-PAGE/CBB staining (Supplementary Fig. S1). Mice were immunized with the purified GST-SSL5, and splenocytes from immunized mice were fused with PAI myeloma cells. Hybridomas were selected with HAT medium and screened by ELISA using purified GST-SSL5 or GST. After repeated limiting dilution, we obtained hybridomas producing anti-SSL5 and anti-GST monoclonal antibodies: clone 30G5C (IgG1/κ) against SSL5 and clone 28A7 against GST (IgG2a/κ). In immunoblotting analysis, anti-SSL5 antibody (30G5C) gave a single band at ∼48 kDa corresponding to GST-SSL5, but did not react with GST (Fig. 1A), whereas anti-GST antibody (28A7) detected both GST (∼25 kDa) and GST-SSL5 (∼48 kDa) (Fig. 1B).

Immunoblotting analysis with
Specificity of anti-SSL5 monoclonal antibody
To analyze the specificity of anti-SSL5 antibody (30G5C), we performed immunoblotting using recombinant His-tagged SSLs (His-SSL1 to SSL14). As shown in Figure 2, the anti-SSL5 antibody reacted strongly with His-SSL5 but not with other His-tagged SSLs, whereas all His-tagged SSLs were detected by anti-6xHistidine antibody at the positions corresponding to their respective molecular weights. We next examined whether the antibodies obtained in this study were applicable to immunoprecipitation. Anti-GST antibody (28A7) conjugated with Dynabeads Protein G precipitated both GST (∼25 kDa) and GST-SSL5 (∼48 kDa) (Fig. 3). By contrast, the immunoprecipitate with anti-SSL5 antibody (30G5C)/Dynabeads Protein G conjugate gave only the GST-SSL5 band (∼48 kDa), as well as two bands for IgG subunits, a heavy chain band (50 kDa) and light chain band (25 kDa). These results indicate that the novel anti-SSL5 monoclonal antibody is highly specific for SSL5 and would be an effective tool for the detection of SSL5 in ELISA, immunoblotting, and immunoprecipitation analyses. We then assessed the domain recognized by anti-SSL5 antibody (30G5C). Truncated forms of SSL5, GST-SSL5Nterm (including an OB-fold domain), and GST-SSL5Cterm (including a β-grasp domain) were expressed in E. coli (Fig. 4A), and purified proteins were analyzed by immunoblotting. The results demonstrated that the anti-SSL5 antibody (30G5C) reacted with GST-SSL5Cterm but not with GST-SSL5Nterm (Fig. 4B). However, deletion of either N-terminal 32 amino acid residues or C-terminal 19 amino acid residues from SSL5Cterm (128–234) decreased the binding to the antibody (Supplementary Fig. S2), suggesting that the flanking regions of the β-grasp domain are important for the anti-SSL5 antibody (30G5C) recognition.

Reactivity of anti-SSL5 antibody (30G5C) with recombinant His-tagged SSL proteins. Recombinant His-tagged SSL proteins (His-SSL1 to 14) expressed in E. coli were separated by SDS-PAGE (12.5% polyacrylamide gel) and analyzed by immunoblotting with anti-SSL5 (30G5C) (upper panel) and anti-6xHistidine (lower panel) antibodies.
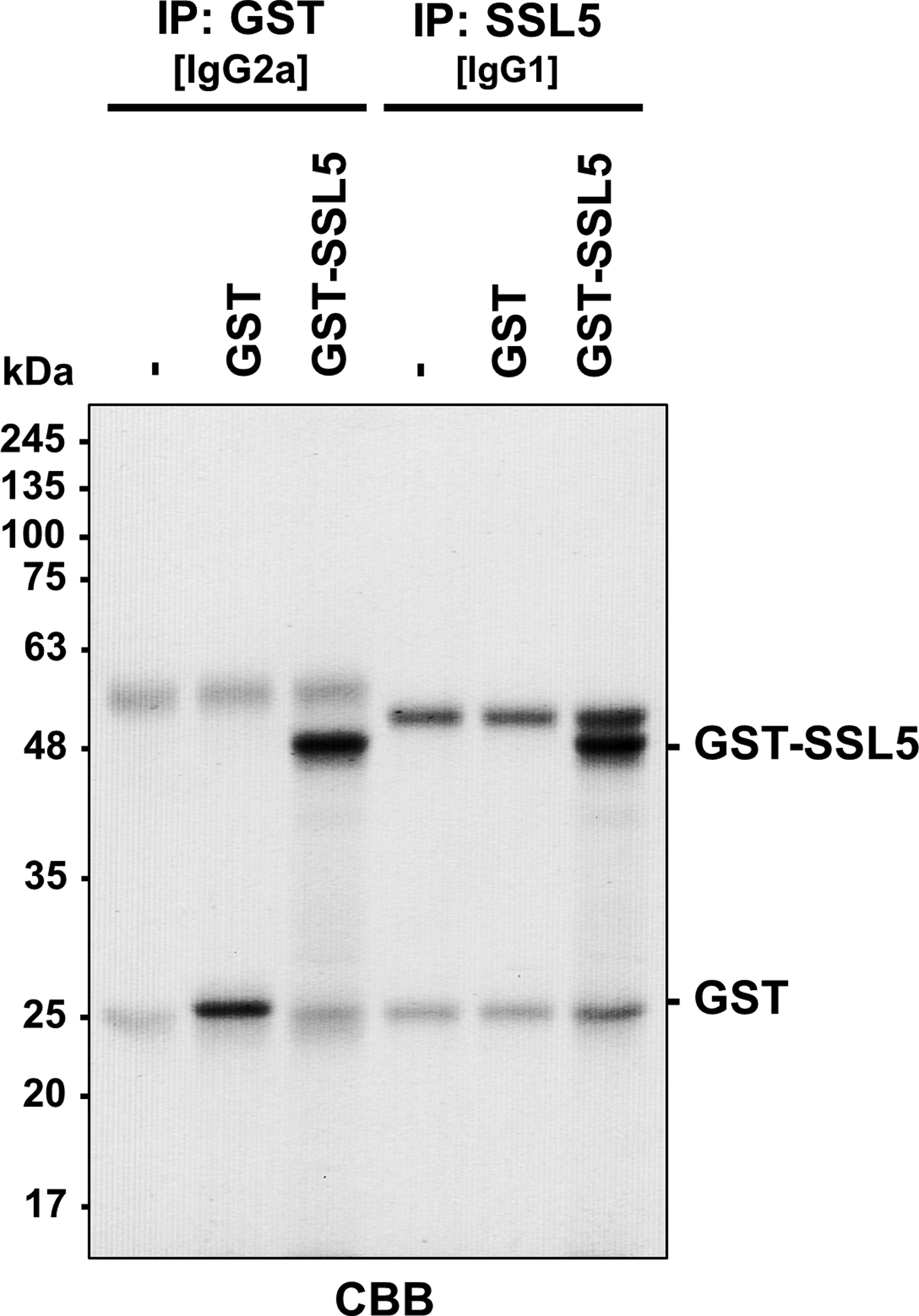
Immunoprecipitation of recombinant GST-SSL5 with anti-SSL5 antibody (30G5C). Recombinant GST and GST-SSL5 were immunoprecipitated with Dynabeads Protein G conjugated with anti-GST (28A7) or anti-SSL5 (30G5C) antibody. The immunoprecipitates were separated by SDS-PAGE (12.5% polyacrylamide gel), and the gel was stained with CBB. The heavy and light chains of IgG gave bands at ∼50 and ∼25 kDa, respectively. CBB, Coomassie Brilliant Blue.
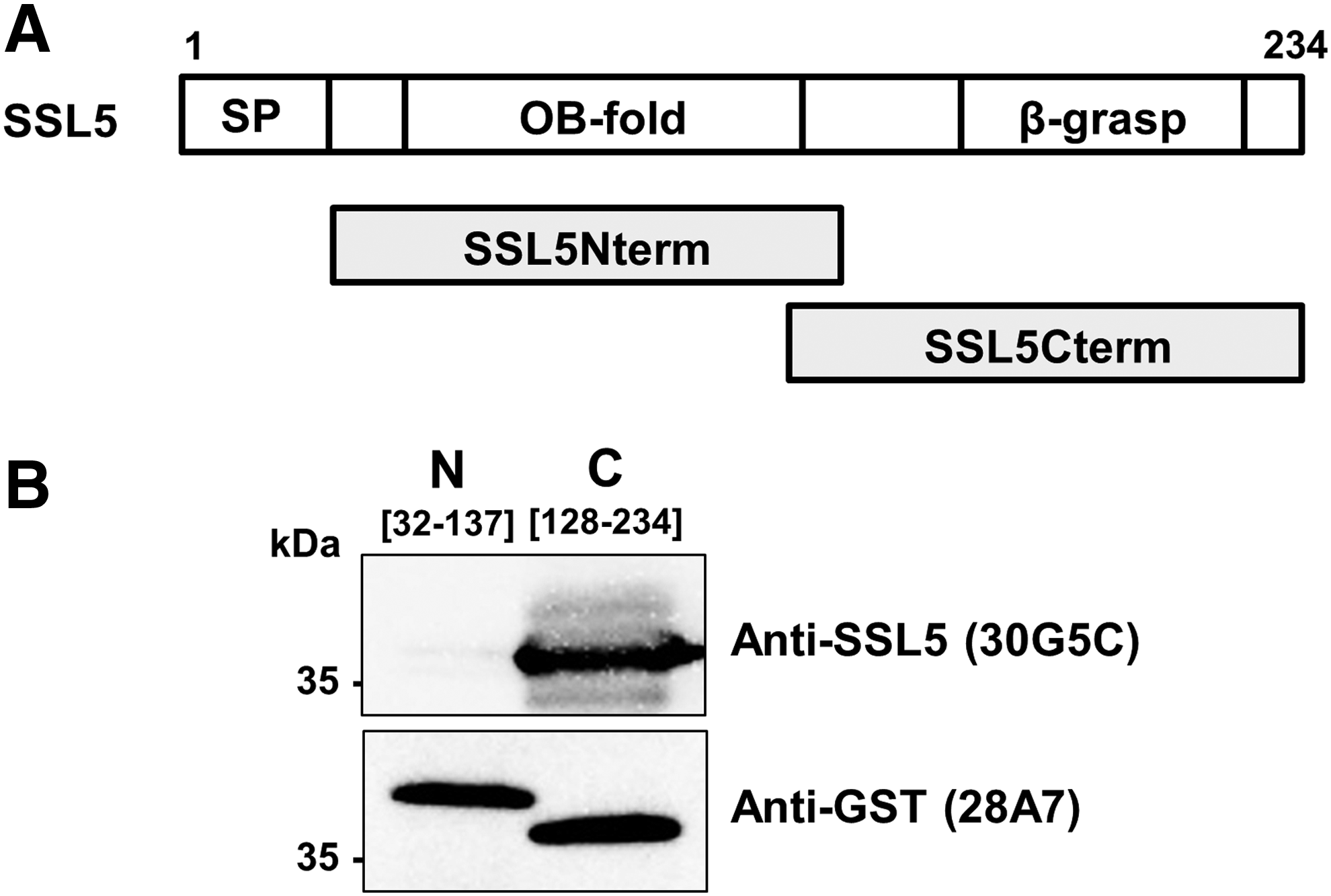
Immunoblotting analysis of the domain recognized by anti-SSL5 antibody (30G5C).
Immunoprecipitation of the SSL5/MMP-9 complex with anti-SSL5 antibody
To examine the effects of anti-SSL5 antibody (30G5C) on the interaction between GST-SSL5 and MMP-9, we purified MMP-9 from conditioned medium of PMA-stimulated THP-1 cells. Electrophoretic analysis of purified MMP-9 gave a single band at ∼92 kDa after CBB staining or immunoblotting (Supplementary Fig. S3). We then performed a pull-down assay using purified MMP-9 and GST-SSL5 conjugated with glutathione agarose in the presence or absence of anti-SSL5 (IgG1/κ). Regardless of the presence or absence of antibody, MMP-9 was precipitated with GST-SSL5/glutathione agarose (Fig. 5). These results indicate that the anti-SSL5 monoclonal antibody (30G5C) does not inhibit the physical association of SSL5 with MMP-9. Finally, we conducted immunoprecipitation of the SSL5/MMP-9 complex with anti-SSL5 antibody (30G5C). When the anti-SSL5 antibody/Dynabeads Protein G conjugate was mixed with MMP-9 in the presence of GST-SSL5, MMP-9 was coprecipitated with GST-SSL5 (Fig. 6). The results of the immunoprecipitation assay indicated the antibody had no inhibitory effect on the binding of GST-SSL5 to MMP-9, and were in good agreement with the results of the pull-down assay (Fig. 5).
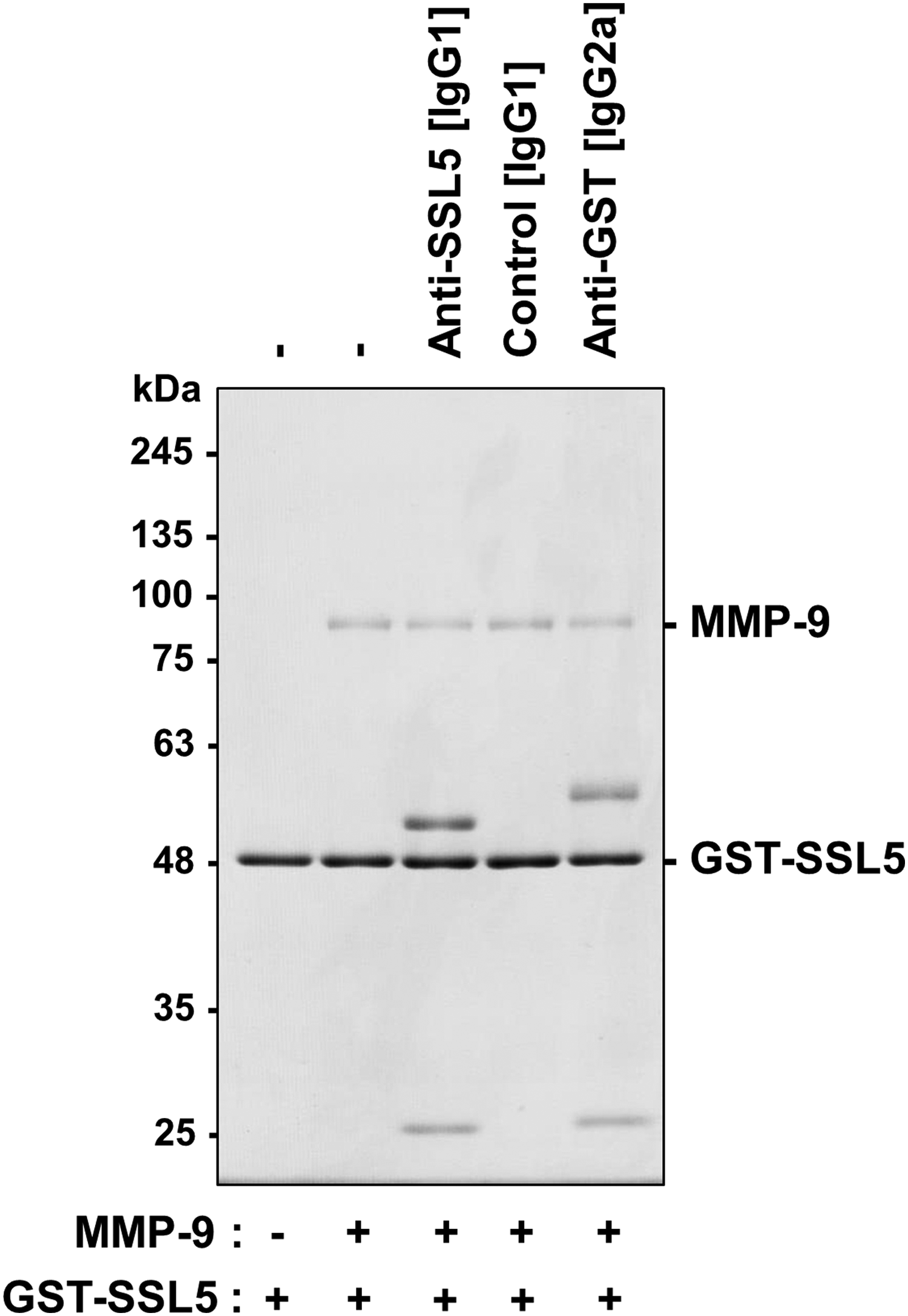
Effects of anti-SSL5 antibody (30G5C) on the pull-down of MMP-9 with GST-SSL5/glutathione agarose. GST-SSL5 (1 μg)-loaded glutathione agarose was incubated with antibody (anti-SSL [IgG1/κ], anti-α3A integrin [isotype-matched control, IgG1/κ] or anti-GST [IgG2a/κ], 1 μg) for 1 hour, and then with purified MMP-9 (1 μg) for 2 hours. Proteins precipitated with GST-SSL5/glutathione agarose were analyzed by SDS-PAGE (10% polyacrylamide gel) followed by CBB staining. MMP-9, matrix metalloproteinase-9.
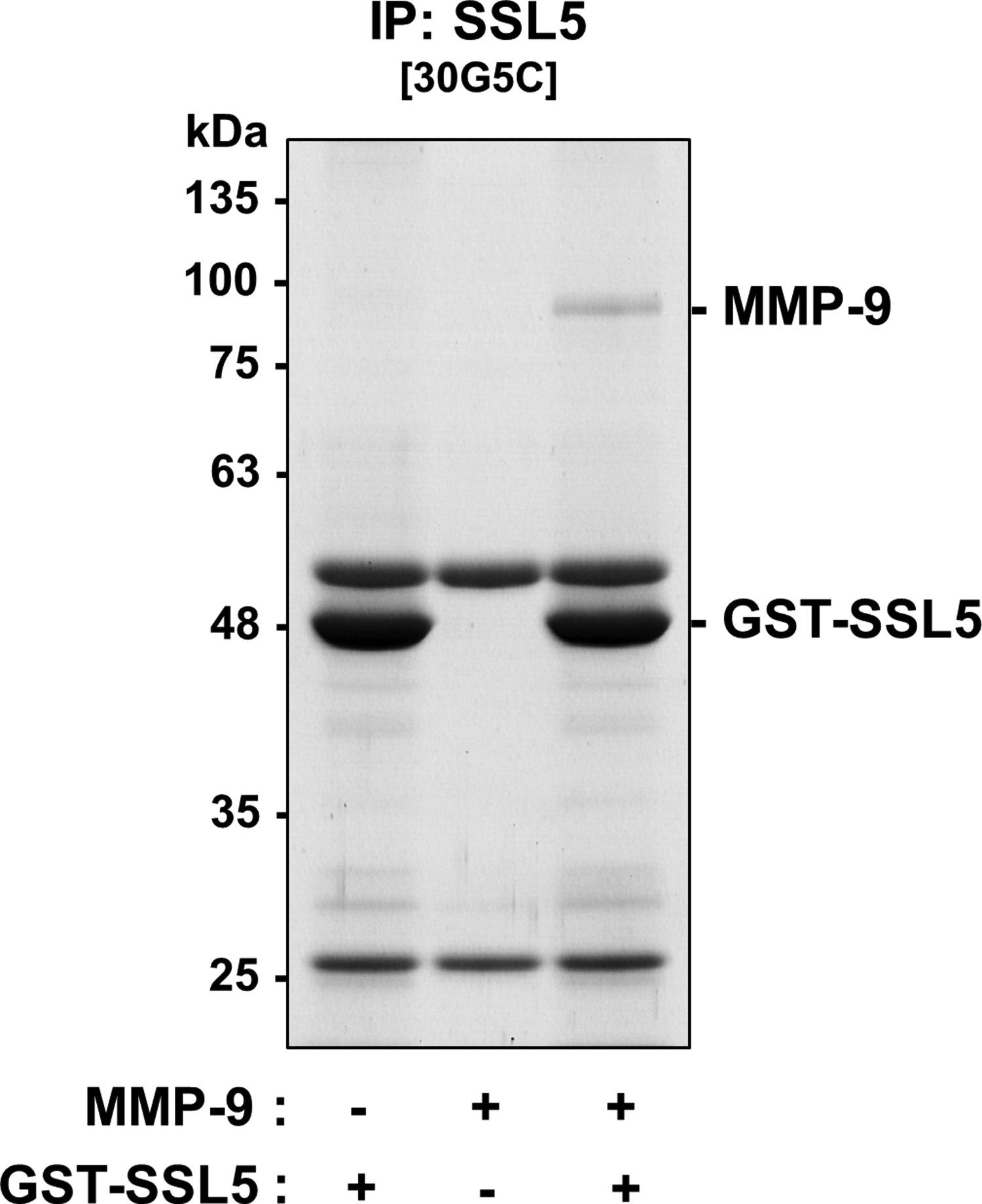
Immunoprecipitation of the GST-SSL5/MMP-9 complex with anti-SSL5 antibody (30G5C). Coimmunoprecipitates with anti-SSL5 antibody (30G5C)-conjugated Dynabeads Protein G were analyzed by SDS-PAGE (10% polyacrylamide gel) followed by CBB staining.
In this study, we developed a monoclonal antibody (30G5C) highly specific for SSL5 without crossreactivity with other members of the SSL exoprotein family and demonstrated that this novel antibody is applicable to ELISA, immunoblotting, and immunoprecipitation (Figs. 1–3). Although a monoclonal antibody reacting with SSL5, SSL7, and SSL11 was reported previously,(24) this antibody (30G5C) seems to be more useful to identify SSL5-producing S. aureus. SSL5 has been considered a virulence factor because it interferes with leukocyte trafficking through its interaction with MMP-9 and PSGL-1.(12,13) Moreover, SSL5 binds to platelets through interaction with cell membrane glycoproteins (GPIb and GPVI) and induces their activation, which causes DIC-related symptoms.(16,24) SSL5 recognizes sialylated glycoproteins (e.g., platelet membrane glycoproteins, FcαRI, PSGL-1, and MMP-9),(14,19,24) and the crucial site for the recognition is thought to be located within the β-grasp motif.(19) We also observed that the binding activity of SSL5 to MMP-9 was abrogated after the introduction of a site-specific mutation within this region (Oku et al., unpublished observation). Although the anti-SSL5 antibody established in this study recognized the β-grasp motif (Fig. 4), the antibody had no inhibitory activity against the interaction between SSL5 and MMP-9 (Fig. 5) and was capable of precipitating the SSL5/MMP-9 complex (Fig. 6). Since SSL5 is one of the potential therapeutic targets for serious or life-threatening diseases associated with staphylococcal infection, this antibody is also suggested to be useful for screening of inhibitors of SSL5/MMP-9 complex formation.
Footnotes
Acknowledgments
This work was supported in part by Grants-in-Aid for Scientific Research from the Ministry of Education, Culture, Sports, Science, and Technology of Japan and by Research Grants from the Shiono Wellness Foundation and the Japan Agency for Medical Research and Development (AMED).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.