
Abstract
We had earlier obtained a murine monoclonal antibody (mAb), termed 5G10, that bound to Salmonella flagellin (SF) and subsequently impaired the latter property of Toll-like receptor 5 (TLR5) signaling activation. Besides interrupting SF-mediated TLR5 activation, mAb 5G10 probably had other potential applications. In this study, we explored multiple functions of 5G10. A short peptide QRVRELAV (designated T5) derived from SF in either terminal of proteins was specifically recognized by 5G10. T5 tag expressed in eukaryotic cell was also detected by 5G10 when analyzed by Western blot, immunofluorescence assay (IFA), and fluorescent-activated cell sorting (FACS). The result of the co-immunoprecipitation assay showed that 5G10 as a bait antibody dragged out the complex of enterovirus 71 (EV71) 2A and mitochondrial antiviral signaling (MAVS) protein. More importantly, 5G10 helped to purify fusion proteins T5-tagged (EV71) 2A and T5-Japanese encephalitis virus NS5 methyltransferase (MTase). Thus, it has been suggested that mAb 5G10 could be useful in several biological applications, including protein identification, location, and affinity purification.
Introduction
T
In an earlier study, we obtained a murine mAb 5G10 against Salmonella flagellin (SF).(6) Besides competing the activity of flagellin on Toll-like receptor 5 (TLR5) signaling, 5G10 was further presumed to specifically recognize a very short linear peptide (residues 90–97 of flagellin, QRVRELAV), which was involved in the flagelin–TLR5 molecular interface.(7) In this study, we verified that 5G10 in combination with peptide QRVRELAV can be used in multiple biological applications, including identification of protein expression, novel protein exploration, and even eukaryocytic protein purification.
Materials and Methods
Cell and plasmids
Human embryonic kidney (HEK) 293T cells were cultured in Dulbecco's modified Eagle's medium (Invitrogen, Carlsbad, CA), supplemented with 10% heat-inactivated fetal bovine serum (FBS) (HyClone, Logan, UT) and penicillin/streptomycin at 37°C in a 5% carbon dioxide humidified atmosphere. Plasmid-containing Japanese encephalitis virus (JEV; SA14-14-2 strain) NS5-MTase gene was a gift from Dr. Cao of Huazhong Agricultural University (HZAU), China.(8) pEF-BO-FLAG-MAVS was provided by Dr. Luo of HZAU, China.(9)
Fluorescent-activated cell sorting analysis
After 48 h transfection with plasmids pIRES-T5-2A, pIRES-2A-T5 or pIRES, 1 × 10 5 293T cells were stained with 5G10 antibody or isotype control 4D1(10) at 4°C for 30 minutes. After washing three times with phosphate-buffered saline containing 1% FBS, the 293T cells were restained with fluorescein (FITC)-labeled goat antimouse immunoglobulin G (IgG) antibody. After washing three times, the 293T cells were analyzed by fluorescent-activated cell sorting (FACS).
Purification of 2A and NS5 methyltransferase
Purified 5G10 antibody (5 mg) was immobilized in a 1 mL HiTrap column (Amersham Pharmacia Biotech) according to the instructions of the manufacturer. The gel in the HiTrap column consisted of highly cross-linked agarose beads, activated by N-hydroxysuccinimide to allow coupling of ligands that contain primary amino groups. Ten 10 cm plates of 293T cells were transfected with pIRES-2A-T5 or pCAGGS-JEV NS5-T5 methyltransferase (MTase)-T5. Forty-eight hours after transfection, cells were lysed and allowed to pass through a column harboring 5G10 agarose beads. The beads were then washed with 80 mL of Tris-buffered saline (pH 7.5) and eluted with 0.1 mg/mL of epitope peptide QRVRELAV.
Immunoprecipitation
Forty-eight hours after transfection with plasmids pIRES-T5-2A, pIRES-2A-T5, or pIRES, cells were lysed with a radioimmunoprecipitation assay (25 mM Tris-HCl buffer [pH 7.4] containing 150 mM sodium chloride, 1% NP-40, 0.25% sodium deoxycholate) containing a protease inhibitor cocktail (Roche, Indianapolis, IN). Cell lyates were incubated with 5G10 antibody or anti-FLAG antibody (Sigma, St. Louis, MO) at 4°C overnight on a rotator in the presence of protein A/G agarose beads (Santa Cruz Biotechnology, Santa Cruz, CA). Immunoprecipitates were subjected to electrophoresis and Western blot analysis.
Results and Discussion
5G10 recognizes either the N- or C-terminal peptide QRVRELAV
We had earlier obtained a murine mAb 5G10 against pathogenic bacteria SF and commensal bacteria K12 flagellin (KF). Interestingly, mAb 5G10 impaired the engagement of flagellin and TLR5 and subsequently resulted in less expression of interleukin 8 and monocyte chemotactic protein 1.(6) The linear epitope (residues 90–97 of SF, QRVRELAV) on flagellin is considered to be crucial for TLR5 activation (Fig. 1A).(7,11) Therefore, we wanted to know whether 5G10 could identify peptide QRVRELAV (T5 tag) when the tag was fused with a target protein.

5G10 recognizes a short epitope QRVRELAV on bacterial flagellin.
To test whether the terminal T5 could be identified, recombinant plasmids were constructed to express residues 1–97 or residues 90–506 of flagellin (Fig. 1B). After being induced by isopropyl-β-
Detecting T5-fused eukaryocytic EV71 2A by 5G10
The T5 tag in either terminal of prokaryotic flagellin could be detected by 5G10. We next set enterovirus 71 (EV71 BrCr strain) nonstructural protein 2A and T5 tag as fusion eukaryocytic protein example to test the binding activity of 5G10 (Fig. 2A). We first constructed two plasmids (designated pIRES-T5-2A and pIRES-2A-T5) that contained a 5′- or 3′-terminal tag in EV71 2A gene. The recombinant plasmids pIRES-T5-2A, pIRES-2A-T5, or pIRES control were transfected to HEK 293T cells. Forty-eight hours later, the transfected cells were harvested and subjected to different analyses, such as IFA, Western blot, and FACS. First, after transfection with the plasmids, cells were fixed and permeated. 5G10 and an isotype antibody control 4D1 were used to detect fusion proteins T5-2A and 2A-T5. The IFA results showed that the fluorescent signal in the transfected cell could be detected by 5G10 but not by the isotype antibody 4D1 (Fig. 2B). Second, the transfected cells were harvested and subjected to SDS-PAGE and the proteins were transferred onto polyvinylidene fluoride membrane. 5G10, but not the isotype control antibody 4D1, detected both fusion proteins T5-2A and 2A-T5 by Western blot assay (Fig. 2C). Third, the transfected cells were subjected to FACS assay after 5G10 dyeing. FITC-labeled goat antimouse IgG was used as the second detection antibody. 5G10 led to numerous transfected cell high fluorescence signal, but not the isotype control antibody (Fig. 2D). Statistically, medium fluorescent intensity in 5G10 dyeing groups was significantly higher than that in the isotype control antibody 4D1 (Fig. 2E) (p < 0.001). Therefore, 5G10 could be used as an alternative antibody to detect expressed eukaryotic fusion protein with a T5 tag.

Detecting T5-fused EV71 2A by 5G10.
Purifying fusion proteins 2A-T5 and NS5 MTase-T5 and dragging out mitochondrial antiviral signaling protein that interacts with EV71 2A protein using 5G10
We also wanted to know whether 5G10 could be used as a bait for protein affinity purification. The 5G10 was first coupled with agarose beads as per the illustration provided by the manufacturer. pIRES-2A-T5 or pCAGGS-JEV (SA14-14-2 strain) NS5-T5 MTase transfected cells were subjected to lyse and passed through a column harboring 5G10 agarose bead. After adequate incubation, the target protein T5-2A or NS5 MTase-T5 was competitively eluted with the peptide QRVRELAV. The 2A-T5 and NS5 MTase-T5 fusion proteins with MW 17 kDa (Fig. 3A) and 65 kDa (Fig. 3B) were eluted, respectively, as shown in the gel of SDS-PAGE. The arrowhead indicates the corresponding proteins 2A-T5 (Fig. 3A) and NS5 MTase-T5 (Fig. 3B). Hence, 5G10 could also be used for affinity protein purification.
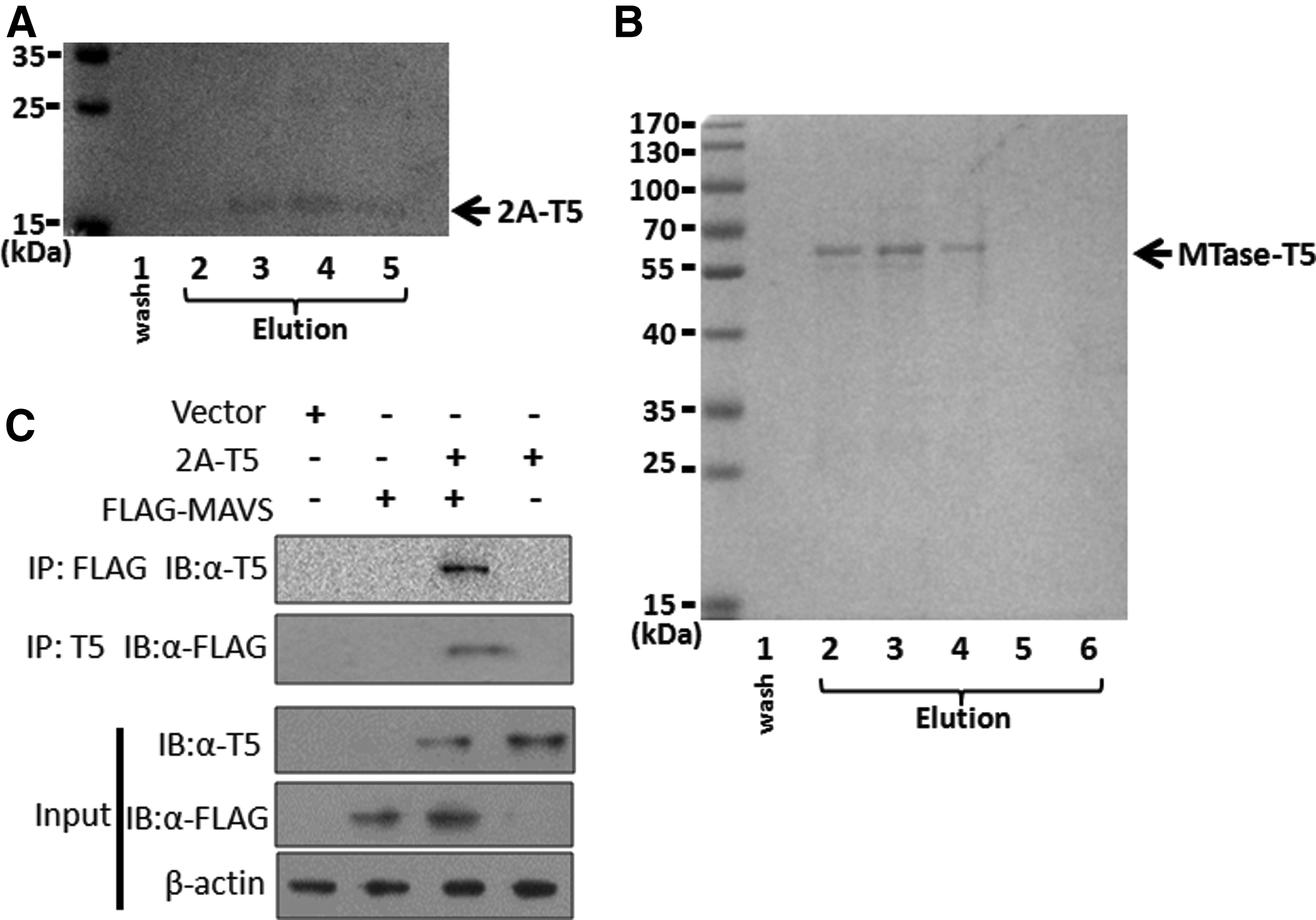
Purification of fusion proteins 2A-T5 and NS5 MTase-T5 and dragging out MAVS protein that interacts with EV71 2A protein using 5G10. 5G10 was coupled on agarose beads according to the manufacturer illustration. pIRES-2A-T5 or pCAGGS-JEV NS5 MTase-T5-transfected cells were lysed and allowed to pass through the column containing agarose beads, and the target protein 2A-T5 or NS5 MTase-T5 was competitively eluted with peptide QRVRELAV finally. The 2A-T5 and NS5 MTase-T5 fusion proteins with MW 17 kDa
As an eight-amino acid short peptide, the protein property of T5 tag could almost not change when fused. Thus, 5G10 could be used as a bait antibody to drag out novel proteins that are interacting with a known protein. As is known, EV71 2A binds to mitochondrial antiviral signaling (MAVS) protein and cleaves it to interrupt interferon α/β production.(12) Here we attempted to assess whether MAVS protein could be pulled out during 2A-T5 baiting by 5G10. pEF-BO-FLAG-MAVS was cotransfected with pIRES-2A-T5 into HEK 293T cells, 48 hours later the cells were subjected to co-immunoprecipitation (Co-IP) assay by 5G10 as a bait antibody. The result showed that 5G10 could drag out the complex of 2A-T5 and its conjugating protein MAVS (Fig. 3C), as could a positive control antibody anti-FLAG. Thus, it demonstrated that 5G10 could be used as a bait antibody for novel protein exploration.
In summary, in this study, we provide solid evidence for applications of mAb 5G10. When fused with the short T5 tag, a target protein could be detected, traced, and even purified by 5G10. Therefore, the murine mAb 5G10 combined with the corresponding T5 tag can be used as potential tools in biological research, including protein location, novel protein exploration, and protein purification.
Footnotes
Acknowledgments
This study was supported by grants from Hubei Provincial Natural Science Foundation of China 2018CFB449, High-Level Scientific Research Foundation for the introduction of talent of Wuhan Institute of Bioengineering (2017KQ01), and National Natural Science Foundation of China (Grant Nos. 81202381 and 81460272).
Author Disclosure Statement
No competing financial interests exist.