
Abstract
Adjuvant can play an important role in vaccine formulation by aiding in the development of a robust immune response. In our hybridoma development work, we typically use both Freund's and AddaVax™ adjuvant regimens for mouse immunizations. While we have repeatedly shown success with our protocols, we continually seek to improve upon the titer and affinity of the serum antibody response. To that end, we evaluated the use of CpG oligodeoxynucleotides (CpG-DNA), a B cell stimulant, in our adjuvant regimens. Mice were immunized using our standard Freund's protocol (Adjulite Complete Freund's Adjuvant for the primary immunization followed by Adjulite Incomplete Freund's Adjuvant (AIFA) for all additional immunizations) or a test protocol using AIFA supplemented with CpG-DNA for all immunizations. A second group of mice were immunized with antigen emulsified in AddaVax adjuvant alone or AddaVax supplemented with CpG-DNA. Our results show a trend toward a higher titer response when CpG-DNA was used with either adjuvant. In addition, AIFA+CpG-DNA mice trended toward a higher relative affinity versus mice immunized using our standard Freund's methodology. Additional antigens will need to be studied to determine whether these observations are limited to the proteins (antigens) studied or whether this is a generalized response to any immunogen.
Introduction
O
The gold standard for use in animal immunizations is Freund's, a water-in-oil adjuvant, which has been used for many years to develop high antibody titers to a variety of antigen types (i.e., proteins, peptides, haptens, and so on) in mice, rats, rabbits, goats, and sheep as well as other species. Freund's adjuvant forms a thick, stable emulsion when mixed with antigen that has been prediluted in saline. Freund's inoculums function via a depot effect that results in a slow, continuous release of antigen to the immune system, usually resulting in a strong, long lasting response.
Freund's adjuvant is available in two formulations: Complete Freund's adjuvant (CFA) comprises heat-killed mycobacteria in paraffin oil, whereas Incomplete Freund's adjuvant (IFA) includes only the oil. While CFA is good at “jump starting” the immune system, the mycobacteria can cause severe inflammation and even abscesses at the site of injection and, therefore, is most commonly used solely for the primary immunization, with IFA being used for all subsequent boosts.
Different companies sell their own formulation of Freund's and we use Adjulite Freund's adjuvant (AFA) from Pacific Immunology for our immunizations. AFA uses a lighter mineral oil versus other Freund's formulations and is thought to be easier on the animals in terms of fewer adverse injection site reactions. The complete version (Adjulite Complete Freund's adjuvant—ACFA) includes 1 mg/mL heat-killed and dried Mycobacterium tuberculosis.
In earlier work with rabbits, we compared AFA to our previously used version of Freund's and found AFA resulted in fewer injection site abscesses, while still yielding a strong response to the immunizing antigen.(2) We have now used AFA in the preponderance of our studies for more than 10 years, with a great deal of success in generating a strong response to a wide variety of antigens in mice, rabbits, sheep, and goats.
An alternate adjuvant is AddaVax™. AddaVax is a squalene-based oil-in-water nanoemulsion based on the formulation for MF59,® which is used in some flu vaccine formulations. The squalene oil in AddaVax is more easily metabolized versus the paraffin oil used in Freund's.(3) AddaVax is classified as an immune potentiator(4) and works by creating an immunocompetent environment that aids in developing a strong, mixed Th1/Th2 immune response. Compared to Freund's, inoculums formulated with AddaVax are more quickly processed from the injection site.(5)
AddaVax is easier to work with than Freund's—depending on the buffer used for the stock immunogen and its concentration, some Freund's inoculum preps do not easily form an emulsion and a great deal of mixing is required. AddaVax preps, on the contrary, only require brief vortexing. Also, due to the nature of Freund's preps (passing back and forth between syringes via a stopcock), some loss of prepared inoculum is common, whereas the loss for AddaVax preps is considerably less.
Unmethylated CpG dinucleotides (CpG-DNA) have been evaluated as an immune stimulator in many human and animal vaccine formulations. Synthetic oligodeoxynucleotides (ODN) contain CpG-DNA motifs which mimic the immunostimulatory effects of bacterial DNA. Human–mouse CpG-DNA has been shown to enhance the immune response in animals. Yang et al.(6) showed CpG-DNA, used alone or with alum adjuvant, could effectively enhance the immune response in mice immunized with a live attenuated oral polio vaccine. In addition, Marcq et al.(7) yielded an improved response in chickens when they removed CFA from the primary immunization and instead used IFA plus CpG-DNA for all injections. The same was demonstrated by Conforti et al.(8,9) in rats and Heifers.
For in vitro work, we use CpG-B DNA (ODN 2006, human/mouse) to stimulate mouse B cells in culture. In addition, we have found it to be beneficial in terms of growth for mouse hybridomas.
To ensure a high titer immune response, we now routinely use a two-pronged adjuvant approach for our mouse immunization programs, using both Freund's and AddaVax adjuvant regimens. The reason here being that, while the adjuvants perform similarly for many antigens, we have observed (unpublished results) that, in some instances, one or the other can yield a significantly higher titer and affinity versus the other. Here, we sought to investigate the application of CpG-DNA in our work. First, we eliminated the use of ACFA at the primary immunization, instead used Adjulite Incomplete Freund's Adjuvant (AIFA) supplemented with CpG-DNA for all immunizations. Second, we tested AddaVax supplemented with CpG-DNA for all immunizations. Based on work cited, we anticipated an enhanced immune response with the addition of CpG-DNA.
Materials and Methods
Mice immunizations
Two separate antigens (each ∼20 KDa molecular weight proteins) sourced directly from the vendor were diluted to the appropriate concentration in saline and mixed 1:1 v/v (volume:volume) with either adjuvant. Adjulite Freund's (Pacific Immunology, Ramona, CA) emulsions were mixed by passing back and forth between two syringes via a three-way stopcock until a thick, stable emulsion was formed. AddaVax (InvivoGen, San Diego, CA) preparations were mixed in a tube by vortexing vigorously for 30 seconds. For selected mice, the inoculum was supplemented with 5 μg/mouse of human–mouse CpG-B DNA (ODN 2006) (HyCult Biotech, Plymouth Meeting, PA). All mice (The Jackson Laboratory, Bar Harbor, ME) were immunized three times, once every 4–6 weeks, with 20 μg antigen.
Freund's mice received a total of 0.2 mL inoculum subcutaneously (s.c.) per immunization, while AddaVax mice received 0.4 mL s.c.; the total inoculum volume for each adjuvant was divided equally between the two axillary and two inguinal regions on each animals' ventral side. Mice were immunized using one of four regimens: (1) primary immunization using ACFA followed by boosters with AIFA (standard Freund's regimen); (2) AIFA+CpG-DNA for all immunizations, including the primary; (3) AddaVax for all immunizations; or (4) AddaVax with CpG-DNA for all immunizations. Animals were bled 2 weeks following the third immunization and sera collected for testing. All protocols were approved by the Abbott Laboratories Institutional Animal Care and Use Committee.
Preparation of rabbit detection antibody
Rabbit polyclonal detection antibodies were prepared for use in screening mouse serum samples. New Zealand White rabbits (Covance, Princeton, NJ) received 200 μg antigen per rabbit (three rabbits per antigen) delivered in Freund's adjuvant (inoculum prepared as indicated above, ACFA for the primary immunization and AIFA for all subsequent immunizations) using the same timing outlined for mice. An equal volume of serum taken from each rabbit 2 weeks following the second or third immunization was pooled (kept separate by antigen system), purified via MabSelect Xtra protein A (GE Healthcare, UK), labeled with biotin using EZ-Link Sulfo-NHS-LC-Biotin (Thermo Scientific, Waltham, MA) per the manufacturer's instructions and qualified by testing for reactivity to solid-phase coated antigen using standard enzyme immunoassay (EIA) protocols.
Mouse sera screening
High protein binding 96-well microtiter plates (BrandTech, Essex, CT) were coated with 100 μL/well sheep anti-mouse IgG, Fc fragment-specific polyclonal antibody (Jackson ImmunoResearch, West Grove, PA) diluted to 2 μg/mL in phosphate-buffered saline (PBS, pH 7.2) and held overnight at room temperature (about 21°C). The antibody was then removed and the plates blocked with 200 μL/well blocking solution (2% w/v bovine serum albumin and 0.5% v/v polysorbate-20 diluted in PBS). Plates were incubated for 30 minutes, the blocking solution removed, and the plates washed.
Next, mouse serum test samples serially diluted in blocking solution were added to the assay plates (100 μL/well), incubated for about 1 hour while shaking, removed, and the plates washed. Antigen (100 μL/well, diluted to 1 μg/mL in blocking solution) was added, incubated for 10 minutes while shaking, removed, and the plates washed. Next, biotin labeled rabbit polyclonal detection antibody (prepared as described herein) was added (100 μL/well, 500 ng/mL in block) to all assay wells, the plates incubated for 30 minutes, the antibody removed, and the plates washed.
Horse radish peroxidase labeled streptavidin (Jackson ImmunoResearch) was then added (200 ng/mL in block, 100 μL/well), incubated for 30 minutes, removed, and the plates washed. Color was developed using 100 μL/well o-phenylenediamine and the signal quenched with 100 μL/well 1 N sulfuric acid. Signal was read at 492 nm. For all wash steps, assay wells were filled 4× with distilled water and aspirated. Shaking was at about 700 rpm.
Relative affinity determination and statistical methods
To determine relative affinity, a single dilution of mouse serum antibody was tested against limiting concentrations of solution phase antigen using the procedure described herein. Data were plotted in KaleidaGraph software (Synergy, Reading, PA) and the Ag50 value (antigen concentration at half-max) calculated using the following calculation: M1 + M2 × M0/(M3 + M0); M1 = 0.05; M2 = 1; and M3 = 10, where M1 = ratio at minimum antigen concentration, M2 = ratio at maximum antigen concentration (max/max), and M3 = approximate antigen concentration at 0 half-max (see Fig. 1, e.g., Ag50 labeled as b/bmax). Data were analyzed for statistical significance using JMP version 8 software (means/analysis of variance and Student's t for comparison).
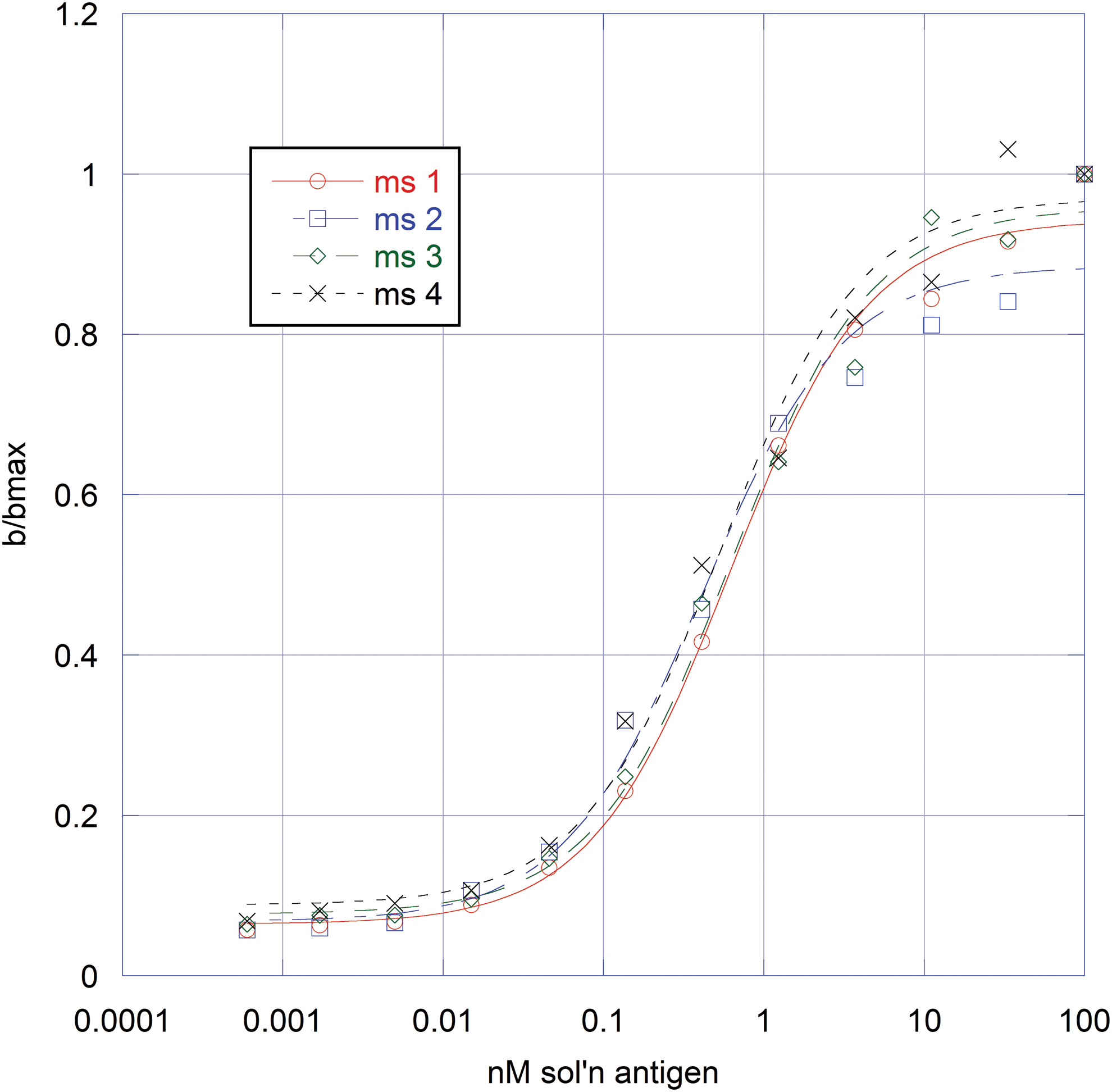
Representative antigen titration curves.
Results
Three New Zealand white rabbits were immunized with each antigen using the standard Freund's regimen. Each animal was bled 10–14 days following the final immunization and the pooled serum purified and labeled with biotin for use in mouse sera screening. Each biotin labeled antibody was tested by microtiter EIA for reactivity to solid phase antigen. Both labeled polyclonal antibodies displayed strong reactivity to their respective antigen and were qualified for use (see, e.g., Fig. 2).

Representative biotin-labeled rabbit antibody titration curve. Reactivity to immunizing antigen and BSA. BSA, bovine serum albumin.
Immune response in mice
In the first study, CAF1/J mice were immunized with antigen using each of the four adjuvant regimens and serum antibody titer and relative affinity determined. Results show that mice immunized using AIFA+DNA yielded a significantly higher titer versus those immunized using the standard Freund's protocol (ACFA/AIFA). These same mice (AIFA+DNA) also yielded a significantly higher relative affinity versus the ACFA/AIFA mice. The highest titer, however, was seen in mice immunized using the two AddaVax regimens, with those immunized with AddaVax+DNA trending toward a slightly higher titer versus those immunized using AddaVax alone. The relative affinity between the two AddaVax groups was identical and about the same as the AIFA+DNA group (Table 1).
(A)–(C) indicates groupings following statistical comparison.
Note: For all tables, titer was measured as average signal (OD @ 492 nm) at 1:40,500 sera dilution. Relative affinity (Ag50) is nM solution antigen. Groups not connected by the same letter are significantly different.
ACFA, Adjulite Complete Freund's Adjuvant; AIFA, Adjulite Incomplete Freund's Adjuvant; OD, optical density at 492 nm.
Building on results from the initial study, additional mice (CAF1/J and SJL/J) were immunized with a second antigen using the standard Freund's, AIFA+DNA, or AddaVax+DNA adjuvant regimens. Animals were immunized and the sera screened using the same methodologies outlined for the initial study. Pooled results from both strains of mice show each adjuvant regimen yielded about the same serum antibody titer, while the AddaVax+DNA and AIFA+DNA groups trended toward higher relative affinity versus the ACFA/AIFA mice (Tables 2 and 3).
No difference between groups in titer or affinity, p = 0.05.
(A) and (B) are groupings following statistical comparison. Groups not connected by the same letter are significantly different.
Comparisons made within each strain, not between strains.
Strain to strain comparison shows that CAF1/J mice yielded a higher titer and relative affinity versus SJL/J mice (Table 4). In addition, combined results from both proteins for ACFA/AIFA, AIFA+DNA, and AddaVax+DNA mice show that those regimens where DNA was included trended toward a higher titer and yielded a significantly higher relative affinity (Table 5).
Groups not connected by the same letter are significantly different.
Groups not connected by the same letter are significantly different.
Discussion
An effective adjuvant is one that can generate a high titer, high-affinity response, regardless of immunogen. Antibodies are regularly raised versus large molecular weight (MW) proteins as well as smaller haptens and peptides and, while there are several adjuvants commonly used in animals which function for most antigens, it can still be difficult to develop the needed response to weakly immunogenic compounds. Identification of an immune system stimulant capable of enhancing the serum antibody titer and affinity would, therefore, be highly beneficial.
CpG-DNA has been extensively evaluated as a component for human and animal vaccines. Its immunostimulatory effects and ease of use make it a prime candidate as a supplement to our current adjuvant regimens. In addition, given the adverse reactions to CFA, identification of an adjuvant regimen does not include the use of CFA. In this study, we used mice serum antibody titer and relative affinity to evaluate the potential benefit to including CpG-DNA in our inoculum formulations.
Results show that the addition of CpG-DNA can, in fact, result in a stronger and higher affinity response. While the use of CpG-DNA with AddaVax showed a trend toward a more potent antigen-specific immune response and similar relative affinity, its use with AIFA in place of our standard ACFA/AIFA regimen showed a clear improvement in strength of response as well as resulting relative affinity. Our results complement those generated by Yang et al.(6) Marcq et al.,(7) and Conforti et al.(8,9) who all showed the addition of CpG-DNA to inoculum resulted in a higher titer response in several animal species.
Results with CpG-DNA in AddaVax adjuvant are encouraging, and in that we have observed some antigens where AddaVax yields a poor response in mice. The addition of CpG-DNA may be what is needed to help sufficiently stimulate B cells in these instances. Just as significantly, we were able to eliminate the use of CFA without losing antibody titer.
A final result worth noting is the significant difference in response to protein 2 from CAF1/J versus SJL/J mice. We commonly use multiple strains of mice with different major histocompatibility complex haplotypes, and, while there are times when we see a strong response from all strains involved, we have found that it is not at all uncommon to find drastic differences in response between various strains of mice.
In summary, the results from our study show a benefit in adding CpG-DNA to the inoculum, and we are continuing this work with additional antigens. The elimination of ACFA needs to be repeated across a broad range of antigen types, including some far less immunogenic such as peptides, smaller MW proteins, and haptens conjugated to carrier proteins, before we can consider completely abandoning the use of ACFA. In addition, we have yet to explore the removal of ACFA from rabbit immunizations.
We have tested AddaVax in rabbits (unpublished work) and found it to be very ineffective and, while the addition of CpG-DNA does enhance antibody titer, the rabbits we tested with AddaVax yielded such a poor response that it is doubtful that the addition of CpG-DNA would be of significant benefit, but certainly worth testing. Finally, in this study, we used CpG-DNA at 5 μg/mouse. The next logical step would be to evaluate its use at higher doses to determine the effect, if any, on antibody titer and affinity.
Footnotes
Acknowledgments
The author thanks B. Tu and A. Scott Muerhoff for their detailed review of the article.
Author Disclosure Statement
No competing financial interests exist.