
Abstract
Collectin LK (CL-LK) is a recently described collectin complex, which upon binding to microbial glycoconjugates, activates the lectin pathway of the complement system, and thereby contributes to the removal of invading microorganisms. The complex consists of the two collectins; Collectin K1 (kidney 1 alias CL-11) and Collectin L1 (liver 1 alias CL-10). At present, most efforts have been made on the characterization of CL-K1, and little is known about the function of CL-L1 and its association with diseases. Deficiency of either of the two collectins is associated with the developmental syndrome 3MC, whereas increased plasma levels of CL-K1 are associated with disseminated intravascular coagulation. Using CL-LK purified from human plasma as an immunogen, we succeed in generating seven monoclonal antibodies (mAbs) with specificity for CL-L1. All seven mAbs recognize both native and recombinant CL-L1. In addition, four of the mAbs were successful in immunohistochemical detection of CL-L1 in human tissues. To our knowledge, these are the first mAbs specific for human native CL-L1 described in the literature, and we expect them to be of great importance in characterizing the function of CL-L1, as well as for the study of CL-L1's association with disease.
Introduction
T
At present, little is known about the function, tissue expression, and the role of CL-L1 in relation to disease; however, deficiency is associated with the developmental syndrome 3MC.(7,8) In the quest for enhanced understanding of the physiological function of CL-L1 and its role in relation to health and diseases, monoclonal antibodies (mAbs) specific for CL-L1 are invaluable tools. To our knowledge, there are only few mAbs specific for human CL-L1 described in the literature and these appear not to recognize native CL-L1.(9) Therefore, we set out to generate CL-L1-specific mAbs using the collectin complex CL-LK as an immunogen.
Materials and Methods
Purification of CL-LK from plasma
Native CL-LK was purified from human plasma by affinity chromatography using a calcium-sensitive mAb against CL-K1 as previously described.(10) CL-LK used for the immunization was further purified by ion exchange chromatography on a 1 mL Source 15Q column (GE Healthcare) equilibrated in 20 mM Tris with 50 mM NaCl and pH 8.0. Bound proteins were eluted with a 50–500 mM NaCl gradient spanning 20 mL.
Expression and purification of recombinant CL-L1 and CL-K1a
Full-length untagged human CL-L1 and human CL-K1a were expressed in CHO DG44 cells using a pOptiVec vector (Invitrogen) as previously described.(4) Recombinant CL-K1a was purified using the same protocol as for the purification of natural CL-LK. Recombinant CL-L1 was purified using the CL-L1-specific mAbs described herein and elution with low pH.
Immunization and hybridomas
For the immunization, purified natural CL-LK was dissolved in tris-buffered saline (TBS) with 5 mM EDTA and mixed with GERBU ADJUVANT P (GERBU Biotechnik, GmbH). Two female NMRI mice were immunized subcutaneously with 20 μg of CL-LK twice, with an interval of 2 weeks, before intravenous immunization 3 days ahead of fusion of the spleen cells. The generation of hybridomas was carried out as previously described and by the principles of Köhler and Milstein.(11,12) Supernatants were screened against natural CL-LK and recombinant CL-K1a coated in enzyme-linked immunosorbent assay (ELISA) wells, and the hybridomas were divided into three groups. Group II comprised hybridomas secreting mAbs that only recognized natural CL-LK and these hybridomas secreted mAbs likely specific for CL-L1, CL-K1d, or a conformational CL-LK epitope. The two other groups of hybridomas secreted antibodies recognizing CL-K1a/d, judged by reactivity in Western blotting of natural CL-LK, or conformational epitopes found on only natural CL-LK, respectively. Recombinant CL-L1 was at the time unavailable for screening purposes, partly due to lack of CL-L1-specific antibodies—later to be used in affinity purification hereof. Uncloned hybridomas were cultured for 1–2 weeks before harvesting supernatant and storage at −80°C. Four milliliters of the supernatants from each of the hybridomas were separately stored at −80°C for screening purposes. The hybridoma cryo library comprised a total of 354 not fully cloned hybridomas.
Screening by Western blotting
A total of 102 hybridomas from group II of the hybridoma collection were screened by Western blotting. Purified natural CL-LK was reduced and separated by 4%–12% Bis-Tris gels (Invitrogen) and electrophoretically transferred to polyvinylidene difluoride membranes (Milipore) using previously described conditions.(13) Hybridoma culture supernatants, containing relevant antibodies, were diluted 1:5, detected with horseradish peroxidase (HRP)-conjugated rabbit anti-mouse Ig (1.3 μg/mL) and developed using carbazol for initial screenings and chemiluminescence (ECL Prime Western Blotting Detection Regent kit; GE Healthcare) for final verification of specificity.
Cloning and purification of anti-CL-L1 mAbs
Hybridomas positive for reactivity with CL-L1 by Western blotting underwent three to six rounds of cloning by limiting dilution. For each round of cloning, the reactivity with CL-L1 was verified by Western blotting. mAbs were purified by Protein A affinity chromatography and bovine IgG absorbed by affinity chromatography, using a column conjugated with rabbit anti-bovine IgG antibodies.
Verification of reactivity
Microwell plates (Maxisorp, NUNC) coated with recombinant CL-K1 or recombinant CL-L1 at a concentration of 2 μg/mL were washed and blocked in TBS with 0.05% emulphogene and incubated with anti-CL-L1 mAbs (20 ng/mL) for 1½ hours. Bound mAbs were detected by HRP-conjugated rabbit anti-mouse IgG (2 μg/mL; Dako) and color was developed with H2O2 and o-phenylenediamine. The assay was performed in either the presence of EDTA (5 mM) or calcium (2 mM).
Immunohistochemistry
Immunohistochemistry was essentially carried out as previously described using paraffin-embedded formalin-fixed human tissue sections and detection of primary antibodies by EnVision+ System HRP-labeled polymer (Dako)/DAB+ (Dako).(14,15) In brief, paraffin-embedded formalin-fixed human tissue sections were deparaffinized and rehydrated with xylene and decreasing concentrations of ethanol. In brief, paraffin-embedded formalin-fixed human tissue sections were deparaffinized and rehydrated with xylene and decreasing concentrations of ethanol. Endogen peroxidase activity was blocked with H2O2, and antigens were retrieved by microwave boiling. Primary antibodies were applied in the following concentrations: mAb 16-1 (0.25 μg/mL), 16-5 (2 μg/mL), 16-12 (1 μg/mL), and 16-13 (2 μg/mL) and mouse mAb anti-chicken IgY (1 μg/mL) as control for 1 hour.
Surface plasmon resonance
Binding characteristics of the mAbs were investigated by SPR on a Biacore 3000 instrument (Biacore, Sweden) with immobilized recombinant CL-L1 on a CM5 chip as described previously in details.(16) In brief, CL-L1 was immobilized on EDC-/NHS-activated flow cells by injecting the CL-L1 (10 μg/mL) in 10 mM acetate pH 5.0 (surface density: 0.028–0.042 pmol/mm 2 ). Ethanolamine, pH 8.5, was used for capping. Running buffer consisted of 10 mM HEPES, 150 mM NaCl2, 5 mM EDTA, and 0.005% Tween-20, pH 7, and mAbs were applied in concentrations ranging from 2.08 to 166 nM. Each sample (40 μL) was injected with a flow rate of 5 μL/min. Regeneration was obtained by two cycles of 100 mM glycine, 5 mM EDTA, 500 mM NaCl, and 0.05% Tween-20, pH 3.0. The apparent dissociation constants were found by fitting the curves to a 1:1 binding model. All experiments were as a minimum conducted as triplicates of duplicates.
Results and Discussion
A collection of hybridomas against CL-LK was generated from immunization of mice with natural human CL-LK; a complex consisting of CL-K1a/d and CL-L1. Group II of this collection (see Materials and Methods section) included hybridomas likely secreting mAbs specific for CL-L1, CL-K1d, or a conformational CL-LK epitope, and these hybridomas underwent several rounds of cloning and screening procedures, including Western blotting, resulting in seven viable hybridomas in the end: Hyb 16-1, 16-3, 16-4, 16-5, 16-7, 16-12, and 16-13, secreting mAbs with specificity for CL-L1. We found that five of seven mAbs by Western blotting reacted with CL-L1 in serum with high specificity (Fig. 1A) and that all seven mAbs recognized both natural CL-LK and recombinant CL-L1 in a calcium-independent manner (Fig. 1B).
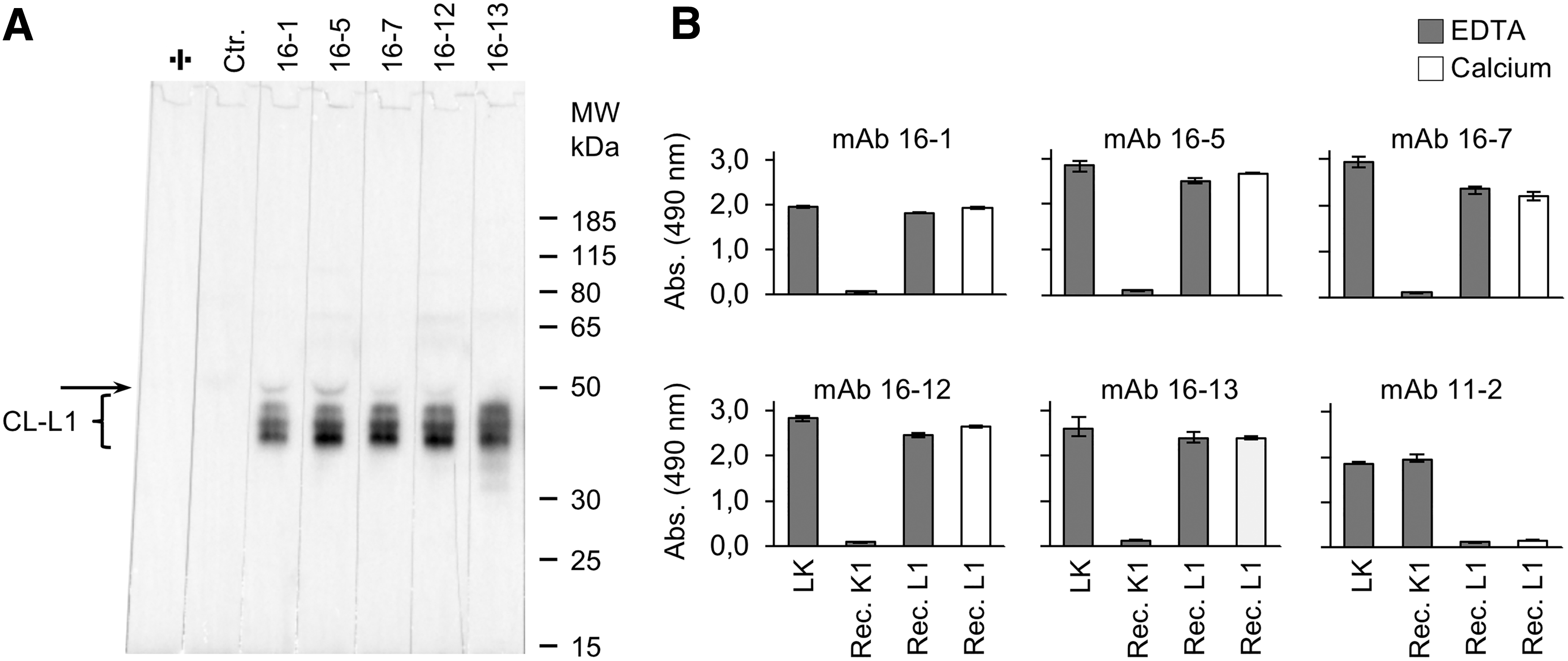
Reactivity of mAbs specific for CL-L1.
Four of the seven mAbs (16-1, 16-5, 16-12, and 16-13) were furthermore successful in immunohistochemical analyses, with varying staining intensities in tissues already known to express CL-L1, that is, the liver and kidney (Fig. 2 and Table 1).(3) In the liver, positive staining was associated with hepatocytes. In the kidney, positive staining was mainly associated with the tubular system. Partly depending on which mAbs that we applied, certain tissues, such as the liver, showed a varying degree of staining. The differences in reactivity on certain tissue sections may result from differential reactivity with different isoforms of CL-L1 or by differential recognition of CL-L1 in the context of CL-LK heteromers versus CL-L1 homomers or small versus large oligomers. It is likely that the immunogen used for the preparation of the mAb contained both CL-L1/-K1 heteromers and homomers.
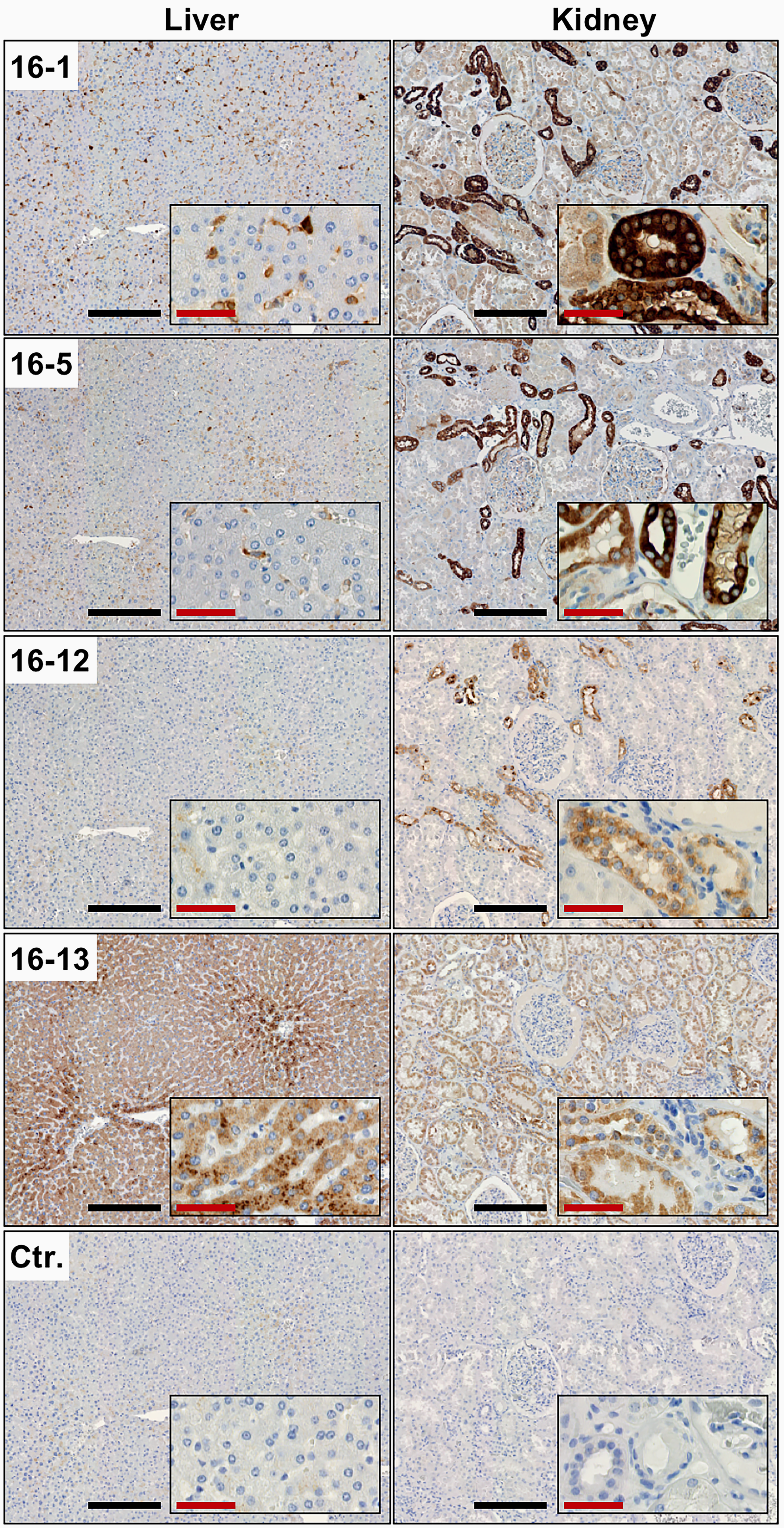
Immunohistochemical analysis using anti-CL-L1 mAbs. Immunohistochemical staining of sections of formalin-fixed and paraffin-embedded human liver and kidney, using four different anti-CL-L1 mAbs (16-1, 16-5, 16-12, and 16-13). A mouse anti-chicken IgY mAb was used as negative control (Ctr.). Four out of the seven anti-CL-L1 mAbs showed reactivity in the immunohistochemical analysis. Black scale bars, 250 μm; red scale bars, 50 μm.
Binding parameters were at least determined by a triplicate run of dilution series of antibody, ranging from 5.0 to 0.08 μg/mL.
IHC, immunohistochemistry; mAb, monoclonal antibody; WB, Western blotting.
By surface plasmon resonance studies we found that the avidity of the intact mAbs, in terms of KD, ranged from 3.1 to 0.63 nM, with mAb 16-3 showing an extreme slow off rate of kd = 3.3 × 10−5/s (Table 1).
Hitherto, levels of CL-L1 in biological fluids have been accessed using polyclonal antibodies raised against CL-L1-derived peptides.(9) The mAbs characterized in this work react also with natural CL-L1 in biological fluids, and a combination of mAb 16-1 and biotin-conjugated mAb 16-13 has proven particularly useful in sandwich ELISA for measurements of CL-L1 levels in biological fluids (not shown).
In summary, we have generated seven mAbs specific for the collectin CL-L1, which is part of the heteromeric complex CL-LK. To our knowledge, this is the first mAbs described in the literature that are specific for human native CL-L1. We expect that the anti-CL-L1-specific mAbs will be extremely useful in, for example, localization studies of CL-L1 and in measuring levels of CL-L1 in association with disease studies, with the purpose of further characterizing the function of CL-L1.
Footnotes
Acknowledgments
This work was in part supported by the A.P. Møller Foundation, Augustinus Foundation, and Dagmar Marshall's Foundation. The authors are thankful to Drs. Karsten Skjødt, Yaseelan Palarasah, and Lars Vitved, University of Southern Denmark for fruitful discussions and helping with the generation of mAbs.
Author Disclosure Statement
No competing financial interests exist.