
Abstract
CHID1 has been recently described as a predictive marker of different malignant tumors. Thus, monoclonal antibodies (mAbs) for CHID1 detection in different human liquids and in tissues are an important tool for the diagnosis of CHID1-positive cancers. However, only few mAbs have been established to date. In this study we describe the generation of a new hybridoma clone 3D4 producing anti-CHID1 antibodies. 3D4 mAb specifically binds human CHID1 and was successfully used in enzyme-linked immunosorbent assay, immunoblotting, immunofluorescence on paraformaldehyde-fixed cells, and in immunohistochemistry of paraffin-embedded tissue specimens. These results indicate that this new anti-CHID1 mAb 3D4 will be useful in the diagnosis of CHID1-related cancers and is a strong tool for both basic and clinical research on chitinase-like proteins.
Introduction
Mammalian chitinase-like proteins (CLPs) are frequently expressed in sites of inflammation or in tumors. Several CLPs are being studied as potential biomarkers of various diseases, including solid tumors (gliomas, prostate, and ovarian tumors). To date there are no clinically approved diagnostic tests for the determination of CLPs in blood or in tissues. Three CLPs are described in humans, these are CHI3L1 (YKL-40, GP39), CHI3L2 (YKL-39), and CHID1 (CHID1, GL008). Chitinase-like proteins are expressed by various cell types in norm and pathology.(1–5) For instance, YKL-40 is expressed by macrophages, neutrophils, epithelial and synovial cells, as well as various tumor cells. YKL-40 may be secreted into extracellular space and in the circulation, which makes it a potential marker for various pathologies. It was demonstrated that this protein stimulates proliferation and invasion of glioma cells(6) and in colorectal tumor cells.(7) YKL-39 is a secretory protein with the highest expression in chondrocytes, synoviocytes, and macrophages. It can be considered as a marker of osteoarthritis in synovial fluid and in serum.(8,9) In pathology, elevated YKL-39 expression is found in patients with Alzheimer's disease and in glioblastomas.(10) The role of YKL-39 in glioblastoma is unclear, but it is suggested to be responsible for proliferative activity of tumor cells and extracellular matrix remodeling.(11) YKL-39 is considered to be a potential glioblastoma marker.
The least studied among these CLPs is CHID1. It was described as an interacting partner of a multifunctional receptor Stabilin-1 in human macrophages.(12) It is expressed in various types of tumors including glioblastomas and also in different inflammatory diseases.(13,14) Recent studies indicate that the expression of CHID1 messenger RNA (mRNA) is significantly downregulated in different brain regions of late onset Alzheimer's disease patients. The authors hypothesize that it is likely to be related to physiological neuronal death.(15) In general, it seems to be feasible to include CLPs in the panels of tumor markers to increase the sensitivity and specificity of existing diagnostic approaches. However, the absence of high-quality antibodies for some CLPs hinders research in this direction.
In this study, we generated a new mouse monoclonal antibody (mAb) that detects CHID1 in enzyme-linked immunosorbent assay (ELISA) and in formalin-fixed paraffin-embedded (FFPE) sections. We characterized this antibody and demonstrated its usefulness for several applications, including immunohistochemistry (IHC) of FFPE sections. Our results suggest that this new anti-CHID1 mAb 3D4 is a powerful tool that can be used in the diagnostics of CHID1-positive cancers and other diseases.
Materials and Methods
Plasmid construction
Total RNA was isolated out of LN229 cells using Qiagen RNeasy Plus Universal Mini Kit (Qiagen, Hilden, Germany) according to the recommendations of the manufacturer. Obtained RNA was used for complementary DNA (cDNA) synthesis using RevertAid Reverse Transcriptase Reverse Transcription Kit (Thermo Fisher Scientific). Obtained cDNA was used to amplify the coding sequence of human CHID1 using polymerase chain reaction (PCR). The following primers were used: forward 5′-ACC TCT CGA GAC CAT GCG GAC ACT CT and reverse 5′-CCT AAA GCT TGA GCA GGT CGT AGA AG. The amplified product was purified out of the agarose gel using Wizard® SV Gel and PCR Clean-Up System (Promega) and cloned into pcDNA3.1(−) myc-His vector between XhoI and HindIII restriction sties. Obtained pcDNA3.1(−) CHID1-Myc-His plasmid was propagated in XL1-Blue Escherichia coli (Evrogen, Moscow, Russia). The integrity of the sequence was controlled by sequencing. Plasmid for transfection was isolated using PureYield Plasmid Midiprep system (Promega).
For generation of a recombinant CHID1, its coding sequence was cloned into pET45b(+) vector between XhoI and HindIII restriction sties. Expression of recombinant protein was carried out in E. coli BL21 DE3 cells.
CHID1 fragments comprising amino acids 1–99, 1–197, and 1–295 were also cloned into pET45b(+) between XhoI and HindIII restriction sties and expressed in E. coli BL21 DE3 cells.
For generation of stably transfected eukaryotic cells, CHID1 coding sequence was cloned into a pLXSN-neo plasmid between EcoRI and XhoI sites, resulting in pLXSN-neo-CHID1-Myc plasmid. The integrity of the sequence was controlled by Sanger sequencing. Plasmid for transfection was isolated using PureYield Plasmid Midiprep system (Promega).
Recombinant protein expression and purification
Expression of recombinant proteins in bacteria was induced in E. coli BL21DE3 with 1 mM IPTG for 4–5 hours at 37°C. Obtained bacteria were lysed using an ultrasound homogenizer. Inclusion bodies were extracted with 5 M urea in PBST for 1 hour at room temperature (RT). Proteins were further solubilized in a lysis buffer, containing 0.5 M arginine, 6 M urea, 20 mM Tris pH 10, 50 mM DTT, 5 mM EDTA, and 10 mM glycine for 12 hours at +4°C. Buffer exchange and refolding were done on a G-25 column with the buffer containing 0.5 M NaCl, 4 M urea, 0.5 M arginine, 1 mM EDTA, 10 mM cysteamine, and 2 mM imidazole, followed by incubation for 36 hours at 4°C. Further protein purification was done on IMAC Sepharose 6 column (GE Healthcare). Elution of CHID1 was effective at 10 mM imidazole. Obtained protein was used for immunization.
Generation of anti-CHID1 antibodies
Recombinant CHID1 protein was mixed with Freund's complete adjuvant (ratio 1:1) and used for immunization. BALB/c mice (16–18 g) were immunized with 30 μg of immunogen per mouse. Subsequent doses were injected with incomplete Freund's adjuvant at 2-week intervals. After two immunizations, popliteal lymph nodes were collected and fused with sp2/0 myeloma cells in a 50% polyethylene glycol 1500 solution (Sigma-Aldrich). The fused cells were plated in 96-well plates in the RPMI medium containing 15% FetalClone serum (HyClone), glutamine, gentamicin, sodium pyruvate, and HAT solution (Invitrogen). After 5 days of incubation at 37°C with 5% CO2 in a humidified environment, culture supernatants were collected and screened for their ability to bind to recombinant protein immobilized on 96-well plates using indirect ELISA. Selected positive hybridoma clones were expanded and subcloned by limiting dilution.
Enzyme-linked immunosorbent assay
Recombinant CHID1 was diluted to 5 μg/mL by phosphate-buffered saline (PBS) and microtiter plate was coated with the protein overnight at 4°C. The plates were washed three times with PBS and blocked with 5% bovine serum albumin (BSA) in PBS at 37°C for 2 hours. The plates were then incubated with antiserum or mAbs. Then the plates were washed four times with PBST (PBS containing 0.05% Tween-20) and incubated with horseradish peroxidase-labeled goat antimouse IgG (1:10,000) at 37°C for 1 hour. Finally, the plates were washed three times with PBST and incubated with freshly prepared TMB with 30% hydrogen peroxide (TMB/H2O2) at 37°C for 10 minutes. The reaction was stopped by 2 M H2SO4. The absorbance was measured with plate spectrophotometer at 450 nm.
mAb subclass identification
The isotype of mAbs was analyzed using the mouse mAb isotyping kit (Genemed, South San Francisco, CA). First, IgA, IgM, IgG1, IgG2a, IgG2b, and IgG3 reagents were diluted by coating buffer (1:1000) and coated overnight at 4°C. The cell culture medium containing mAb was added onto plates and incubated at 37°C for 1 hour. The following procedure was the same as that described in the indirect ELISA.
Cell culture and transfection
Human GP293 and LN229 cells were maintained in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal calf serum (FCS). GP-293 cells were cotransfected with 1.2 μg retroviral vector pLXSN-neo-CHID1-Myc and 0.8 μg pVSVG (Clontech) using Lipofectamine LTX (Thermo Fisher Scientific) according to the manufacturer's protocol. A total of 24, 48, and 72 hours after transfection, virus-containing medium was applied to 50% confluent LN229 cells in the presence of 8 μg/mL Polybrene (Sigma). Infected cells were selected in 1.2 mg/mL G-418-containing medium (Roche) for 7 days.
Immunoblotting
Cells were lysed with a cell lysis buffer RIPA (50 mM Tris-HCl [pH: 7.5], 150 mM NaCl, 2 mM EDTA, 1% Triton X-100, 0.1% sodium deoxycholate, and 1/25 protease inhibitor cocktail [Roche]). After incubating on a shaker for 20 minutes at 4°C, the samples were centrifuged at 16,400 ×g for 10 minutes. The supernatant was collected, and protein concentrations of the lysates were determined by bicinchoninic acid assay (Bio-Rad). Equal amounts of protein from each cell lysate (30 μg per lane) were subjected to sodium dodecyl sulfate polyacrylamide gel electrophoresis. Separated proteins were transferred to a nitrocellulose membrane (Bio-Rad) and the membrane was blocked in Tween 20-tris buffered saline (TTBS; 10 mM Tris-HCl pH 7.6, 0.15 M NaCl, 0.05% Tween20) containing 5% nonfat dry milk (Bio-Rad) for 1 hour, followed by incubation with primary antibodies (anti-CHID1 1.2 mg/mL, 1:1000; anti-Myc-tag 1:1000, No. 2272; Cell Signaling) at 4°C overnight. After washing, the membrane was incubated with horseradish peroxidase-conjugated secondary antibodies (antimouse 1:100,000, antirabbit 1:100,000) for 1 hour at RT. Chemiluminescent detection was carried out with the Novex ECL detection system (Invitrogen) according to the manufacturer's instructions.
Immunofluorescence
To detect CHID1 protein in the cytoplasm, LN229-CHID1 cells were seeded on slides in 2 × 104 cells per well and after 24 hours they were fixed with 3.7% paraformaldehyde in PBS at RT for 15 minutes and permeabilized with 0.1% Triton X-100 in PBS at RT for 3 minutes and then incubated with primary mAb (mouse anti-CHID1 1.2 mg/mL, 1:100; rabbit anti-Myc-tag 1:100, No. 2272; Cell Signaling) at RT for 1 hour. Cells were then stained with the Cy3-conjugated antimouse antibody (1.5 mg/mL, 1:500; Jackson ImmunoResearch) and Alexa488-conjugated antirabbit antibody (1.5 mg/mL, 1:500; Jackson ImmunoResearch) at RT for 1 hour. Images were captured by a fluorescent microscope (Olympus BX53, Tokyo, Japan).
IHC on FFPE section
Immunohistochemical staining was performed on human prostate cancer tissues. Endogenous hydrogen peroxidase was quenched with 3% H2O2 in PBS at RT for 7 minutes. Paraffin-embedded sections (4 mm thick) were deparaffinized and then treated with 10 mM citrate buffer (pH 6.0) at 110°C for 10 minutes by Decloaking Chamber (Biocare Medical). Sections were incubated with primary mAb at 5 μg/mL in PBS at RT for 1 hour and with Poly-HRP DAB Kit (Ms+Rb; Genemed) according to the manufacturer's instructions. Sections were counterstained lightly with Mayer's hematoxylin. Images were captured by light microscopy (Olympus BX53).
Results and Discussion
For generation of anti-CHID1 mAbs, full length recombinant protein was used as immunogen. We obtained 17 hybridoma clones producing IgGs that recognize full length CHID1 antigen in ELISA. One of selected clones was specific to CHID1 and did not bind YKL-40 and YKL-39. ELISA showed OD450 of 3.06 for binding to CHID1, and OD450 of 0.04 and 0.23 for binding to YKL-39 and YKL-40, respectively. The isotype of CHID1 3D4 antibody is IgG1.
To confirm the specificity of the obtained 3D4 antibody, LN229 cells stably transfected with pLXSN-neo-CHID1-Myc were used for Western blotting with 3D4 antibody and anti-Myc antibody. It was found that 3D4 antibody detects protein of the same molecular weight as anti-Myc antibody (Fig. 1A, B) in the lysates of pLXSN-neo-CHID1-Myc-transfected cells. In mock-transfected cells, a faint band with a molecular weight slightly lower than that of CHID1-Myc was detected with 3D4 antibody, most likely corresponding to the endogenous protein. No additional protein was detected in mock-transfected cells using anti-Myc antibody. These data demonstrate that the newly developed CHID1 mAbs are highly specific for detection of CHID1 without cross-reaction to other proteins.

Western blot analysis of CHID1 in LN229 cells constitutively transfected with pLXSN-neo-CHID1-Myc or empty vector with 3D4 (1:1000) antibody
To determine the binding region on novel antibodies, three fragments of CHID1 were expressed in E. coli BL21DE3 (Fig. 1C, D). Western blot analysis of CHID1 fragments with CHID1 3D4 antibody revealed that it binds in the region 99 to 197 AA of CHID1 (Fig. 1C).
To further characterize anti-CHID1 antibody, 3D4 immunofluorescense was used. LN229 cells stably transfected with pLXSN-neo-CHID1-Myc were fixed with paraformaldehyde and stained with anti-CHID1 3D4 antibody and anti-Myc antibody using immunofluorescence to confirm the identity of the protein detected by CHID1 3D4 (Fig. 2). Complete overlap of anti-Myc and CHID1 3D4 staining was observed (Fig. 2D), indicating that 3D4 recognizes recombinant Myc-tagged CHID1. Staining of CHID1 with 3D4 antibody was revealed in LN229 cells nuclear structures, similar to that observed in H1299 cells overexpressing FLAG-tagged CHID1,(12) at the same time it differs from diffuse nuclear staining observed in U-2 OS, A-431, and U-251 MG cells,(16) which can be explained by the differences in cell lines and the presence or absence of necessary sorting receptors.

Specificity of 3D4 mAbs for CHID1 protein was determined by immunofluorescence. LN229-CHID1-Myc cells were plated on cover slips, fixed, permeabilized, and stained with either anti-Myc antibodies used at 1:100 dilution
To test whether 3D4 antibodies are suitable for IHC on FFPE tissues, we have selected tissues documented in ProteinAtlas expression of CHID1. Tissue sections of brain, glioblastoma, colon, and pancreas were stained with 3D4 mAb (Fig. 3A–D). In cerebral cortex, strong cytoplasmic and membranous staining of neuronal cells was observed. Glial cells showed moderate to weak cytoplasmic/membranous staining. No staining of neuropil was found (Fig. 3A). In glioblastoma WHO GIV, moderate to strong membranous–cytoplasmic staining of the tumor cells was observed together with weak staining of the neuropil (Fig. 3B). In large intestine, strong predominantly cytoplasmic staining of the glandular epithelium was observed. Lymphoid cells in mucosal stromal tissue showed moderate to strong membrane–cytoplasmic staining (Fig. 3C). In pancreas, strong cytoplasmic–membranous staining of the exocrine glandular cells and moderate predominantly cytoplasmic staining of the Islets of Langerhans were observed (Fig. 3D). These results indicate that mAb 3D4 can be successfully used to detect CHID1 in FFPE sections. Analysis of CHID1 expression in THP-1 revealed predominantly cytoplasmic staining, which corresponds to results obtained in tissues.
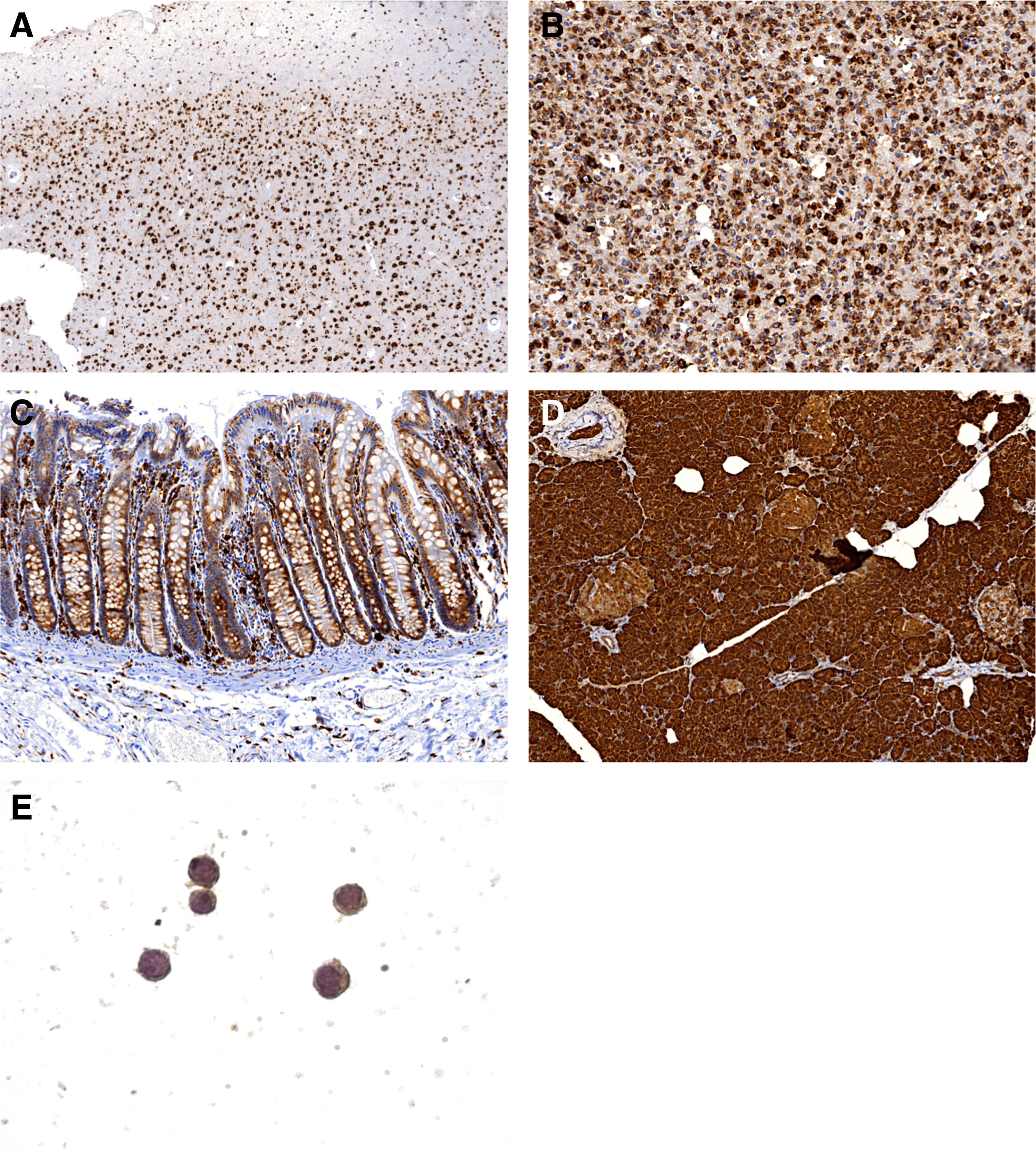
Tissue sections of brain, glioblastoma, colon, and pancreas were stained with 3D4 mAb. In cerebral cortex strong cytoplasmic and membranous staining of neuronal cells was observed. Glial cells showed moderate to weak cytoplasmic/membranous staining. No staining of neuropil was found, magnification 50×
In summary, we describe the development and characterization of a novel anti-CHID1 mAb 3D4 suitable for ELISA, Western blotting, IHC on FFPE sections, and immunofluorescence. The reactivity of this antibody was verified for various types of samples: purified recombinant protein, transfected cells, and different normal and cancer tissues. Thus, our results demonstrate that mAb 3D4 is a mAb against human CHID1 available for IHC of paraffin-embedded sections and suggest that this antibody will be a powerful tool for the study and diagnosis of CHID1-positive cancers and other diseases.
Footnotes
Acknowledgment
This work is supported by Russian Foundation for Basic Research, project 17-04-01857 (A.G.).
Author Disclosure Statement
No competing financial interests exist.