
Abstract
Podoplanin (PDPN), also known as T1alpha, has been used as a lung type I alveolar cell marker in the pathophysiological condition. Although we have established several monoclonal antibodies (mAbs) against mammalian PDPNs, mAbs against tiger PDPN (tigPDPN), which are useful for immunohistochemical analysis, remain to be developed. In this study, we immunized mice with tigPDPN-overexpressing Chinese hamster ovary (CHO)-K1 cells (CHO/tigPDPN) and screened hybridomas producing mAbs against tigPDPN using flow cytometry. One of the mAbs, PMab-231 (IgG2a, kappa), specifically detected CHO/tigPDPN cells using flow cytometry as well as recognized tigPDPN protein using western blotting. In addition, PMab-231 was found to cross-react with cat PDPN (cPDPN). The dissociation constants (KD) of PMab-231 for CHO/tigPDPN and CHO/cPDPN cells were determined to be 1.2 × 10−8 and 1.9 × 10−8, respectively, indicating moderate affinity for CHO/tigPDPN and CHO/cPDPN cells. PMab-231 stained type I alveolar cells of the feline lungs and podocytes of the feline kidneys using immunohistochemistry. Our findings suggest the potential usefulness of PMab-231 for the functional analyses of tigPDPN and cPDPN.
Introduction
Podoplanin (PDPN), also known as a T1alpha, is a type I transmembrane sialoglycoprotein that is expressed in several normal cells, including type I alveolar cells.(1) Therefore, PDPN is extremely useful for distinguishing type I alveolar cells from type II alveolar cells in pathological diagnosis.(2) We previously reported that C-type lectin-like receptor-2 (CLEC-2) is an endogenous receptor of PDPN.(3,4) Importantly, the PDPN/CLEC-2 interaction facilitates embryonic blood/lymphatic vessel separation.(5) PDPN/CLEC-2 interaction also plays an essential role in murine lung development.(2)
The expression of human PDPN (hPDPN) has been reported in several malignant tumors, including brain tumors,(6) oral squamous cell carcinomas,(7) lung cancers,(8) esophageal cancers,(9) malignant mesotheliomas,(10) osteosarcomas,(11) chondrosarcomas,(12) and testicular tumors.(13) The hPDPN expression is associated with cancer metastasis and malignant progression.(4,6,14) Till date, we have developed monoclonal antibodies (mAbs) against human,(15) mouse,(15) rat,(16) rabbit,(17) bovine,(18) dog,(19) cat,(20) pig,(21) and horse(22) PDPNs. However, anti-tiger PDPN mAbs have not been reported. In this study, we immunized mice with CHO/tigPDPN cells and established hybridomas that could produce mAbs against tigPDPN.
Materials and Methods
Cell lines and animals
P3X63Ag8U.1 (P3U1) and CHO-K1 cells were obtained from the American Type Culture Collection (ATCC, Manassas, VA). The tigPDPN bearing an N-terminal LP tag (LP-tigPDPN) was inserted into a pCAG-Neo vector (FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan).(23) The LP tag comprises 16 amino acids (NSVTGIRIEDLPTSES). The CHO-K1 cells were transfected with pCAG-Neo/LP-tigPDPN using Lipofectamine LTX with Plus Reagent (Thermo Fisher Scientific, Inc., Waltham, MA). Stable transfectants were selected by limiting dilution and then cultivated in a medium containing 0.5 mg/mL of G418 (Nacalai Tesque, Inc., Kyoto, Japan). The P3U1, CHO-K1, CHO/tigPDPN, CHO/cat PDPN (cPDPN),(20) CHO/hPDPN,(24) CHO/mouse PDPN (mPDPN),(24) CHO/rat PDPN (rPDPN),(16) CHO/rabbit PDPN (rabPDPN),(17) CHO/dog PDPN (dPDPN),(19) CHO/bovine PDPN (bovPDPN),(18) CHO/pig PDPN (pPDPN),(21) CHO/horse PDPN (horPDPN),(25) CHO/alpaca PDPN (aPDPN),(26) CHO/bear PDPN (bPDPN),(21) CHO/goat PDPN (gPDPN),(27) CHO/sheep PDPN (sPDPN),(28) CHO/Tasmanian devil (tasPDPN),(21) and CHO/whale PDPN (wPDPN)(21) were cultured in Roswell Park Memorial Institute (RPMI) 1640 medium (Nacalai Tesque, Inc.) supplemented with 10% heat-inactivated fetal bovine serum (Thermo Fisher Scientific, Inc.), 100 U/mL penicillin, 100 μg/mL streptomycin, and 25 μg/mL amphotericin B (Nacalai Tesque, Inc.). The cells were grown at 37°C in a humidified environment with 5% CO2 and 95% air atmosphere.
Hybridoma production
Female BALB/c mice (aged 6 weeks) were purchased from CLEA Japan (Tokyo, Japan). The animals were housed under specific pathogen-free conditions. The Animal Care and Use Committee of Tohoku University approved all animal experiments. We employed a Cell-Based Immunization and Screening (CBIS) method(20,29–31) to develop sensitive and specific mAbs against tigPDPN. In brief, two BALB/c mice were immunized with CHO/tigPDPN cells (1 × 108), which were intraperitoneally (i.p.) administered with Imject Alum (Thermo Fisher Scientific, Inc.). The procedure included three additional immunizations, followed by a final booster injection i.p. administered 2 days before the harvesting of the spleen cells, resulting in total five immunizations. These spleen cells were subsequently fused with P3U1 cells using PEG1500 (Roche Diagnostics, Indianapolis, IN), and the hybridomas were grown in RPMI medium supplemented with hypoxanthine, aminopterin, and thymidine for selection purpose (Thermo Fisher Scientific, Inc.). The culture supernatants were screened using flow cytometry.
Flow cytometry
The cells were harvested after a brief exposure to 0.25% trypsin/1 mM ethylenediaminetetraacetic acid (Nacalai Tesque, Inc.). The cells were washed with 0.1% bovine serum albumin/phosphate-buffered saline (PBS) and then treated with primary mAbs for 30 minutes at 4°C. Next, the cells were treated with Alexa Fluor 488-conjugated anti-mouse IgG (1:2000; Cell Signaling Technology, Inc., Danvers, MA) or Oregon Green anti-rat IgG (1:2000; Thermo Fisher Scientific, Inc.). Fluorescence data were collected using the SA3800 Cell Analyzers (Sony Corp., Tokyo, Japan).
Determination of binding affinity using flow cytometry
CHO/tigPDPN was suspended in 100 μL of serially diluted PMab-231, followed by the addition of Alexa Fluor 488-conjugated anti-mouse IgG (1:200; Cell Signaling Technology, Inc.). Fluorescence data were collected using the EC800 Cell Analyzer (Sony Corp.). The dissociation constant (KD) was obtained by fitting the binding isotherms to built-in one-site binding models in GraphPad PRISM 6 (GraphPad Software, Inc., La Jolla, CA).
Western blotting
Cell lysates (10 μg) were boiled in sodium dodecyl sulfate sample buffer (Nacalai Tesque, Inc.). The proteins were subjected to electrophoresis on 5%–20% polyacrylamide gels (Nacalai Tesque, Inc.) and subsequently transferred onto a polyvinylidene difluoride membrane (Merck KGaA, Darmstadt, Germany). After blocking with 4% skim milk (Nacalai Tesque, Inc.), each membrane was incubated with primary mAbs, such as 1 μg/mL PMab-231, 10 μg/mL anti-LP tag (LpMab-17), 1 μg/mL anti-MAP tag (PMab-1), or 1 μg/mL anti-β-actin (AC-15; Sigma-Aldrich Corp., St. Louis, MO), and subsequently with peroxidase-conjugated anti-mouse IgG (1:1000; Agilent Technologies, Santa Clara, CA) or anti-rat IgG (1:10,000; Sigma-Aldrich Corp.). The bands were visualized with the ImmunoStar LD (FUJIFILM Wako Pure Chemical Corporation) using the Sayaca-Imager (DRC Co. Ltd., Tokyo, Japan).
Immunohistochemical analyses
The normal feline tissues were purchased from Zyagen (San Diego, CA). Paraffin-embedded tissue sections (4 μm in diameter) were directly autoclaved in citrate buffer (pH 6.0; Nichirei Biosciences, Inc., Tokyo, Japan) for 20 minutes. These tissue sections were then blocked with the SuperBlock T20 (PBS) Blocking Buffer (Thermo Fisher Scientific, Inc.), incubated with PMab-231 (5 μg/mL) for 1 hour at room temperature, and finally treated using the Envision+ Kit (Agilent Technologies, Inc.) for 30 minutes. Color was developed with 3,3′-diaminobenzidine tetrahydrochloride (Agilent Technologies, Inc.) for 2 minutes, and counterstaining was performed with hematoxylin (FUJIFILM Wako Pure Chemical Corporation).
Results and Discussion
In this study, we employed the CBIS method to develop sensitive and specific mAbs against tigPDPN to facilitate the immunohistochemical analysis of paraffin-embedded tissue sections. Two mice were immunized with CHO/tigPDPN cells by following a screening procedure (Fig. 1). The developed hybridomas were seeded into 96-well plates and cultivated for 7 days. Wells positive for CHO/tigPDPN and negative for CHO-K1 were selected using flow cytometry. Screening identified strong signals against CHO/tigPDPN cells and weak or no signals against CHO-K1 cells in 18 of 958 wells (1.9%). One of the six clones, PMab-231 (IgG2a, kappa), was finally selected using immunohistochemistry against paraffin-embedded feline normal tissue sections because tigPDPN and cPDPN shared 99% homology.(23)
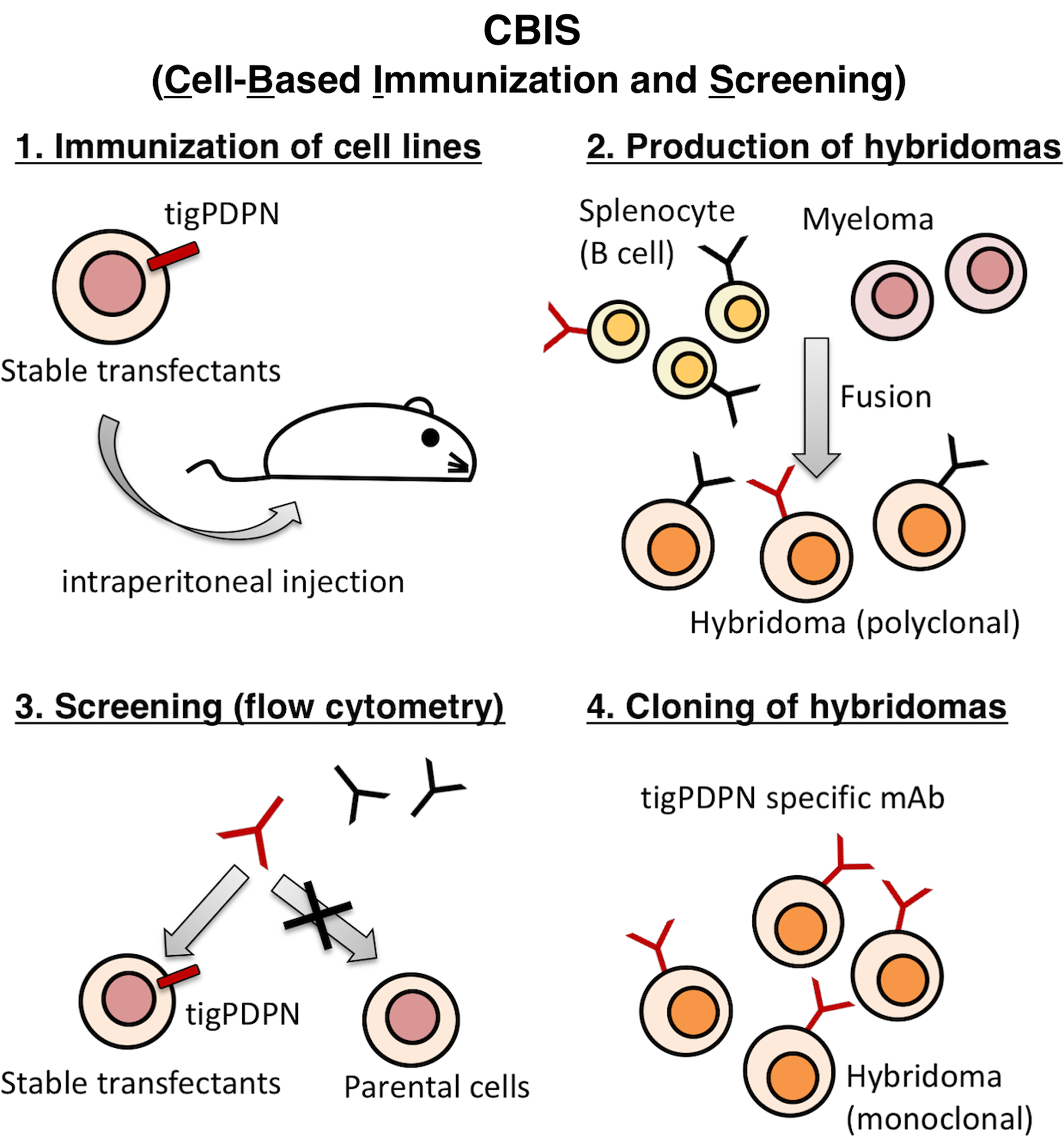
Schematic illustration of CBIS method. Stable transfectants expressing tigPDPN used as an immunogen with no purification procedure. The selection of hybridomas secreting specific mAbs performed by flow cytometry using parental and transfectant cells. CBIS, Cell-Based Immunization and Screening; mAbs, monoclonal antibodies; tigPDPN, tiger podoplanin.
PMab-231 recognized CHO/tigPDPN, but showed no reaction with CHO-K1, as assessed using flow cytometry (Fig. 2). LpMab-17, an anti-LP tag mAb, detected CHO/tigPDPN. PMab-231 cross-reacted with cPDPN and horPDPN, but did not react with human, mouse, rat, rabbit, dog, bovine, pig, alpaca, bear, goat, sheep, Tasmanian devil, or whale PDPNs (Fig. 3).
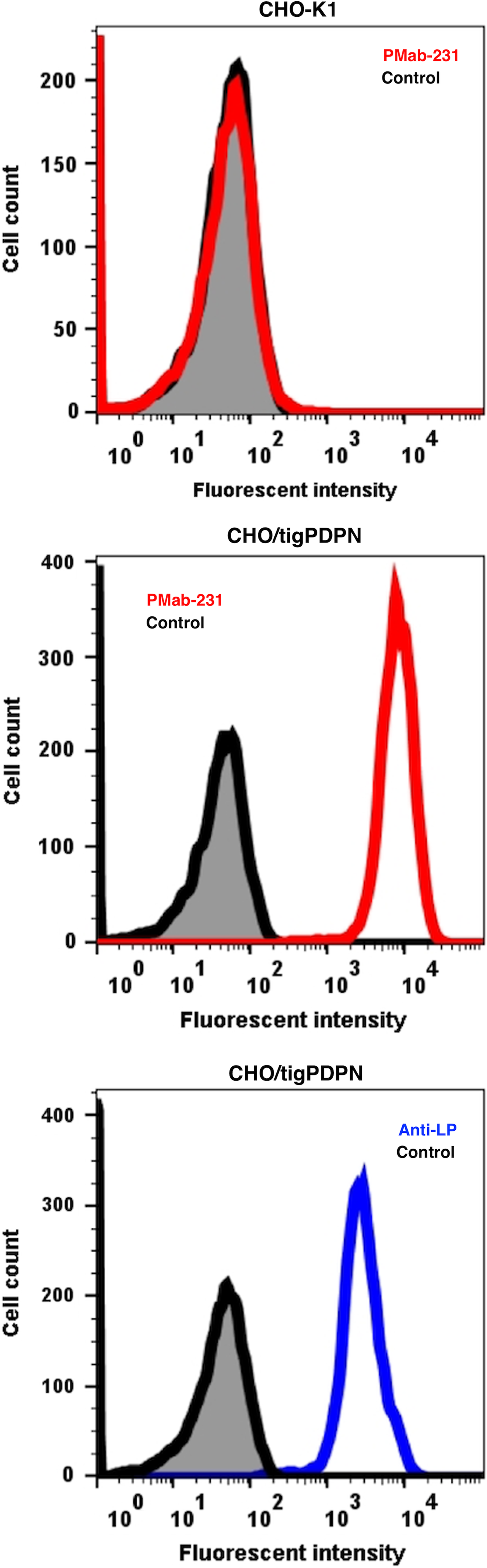
Detection of tigPDPN by flow cytometry using PMab-231. CHO/tigPDPN and CHO-K1 cells were treated with PMab-231 (red line) or anti-LP tag (LpMab-17; blue line) at a concentration of 1 μg/mL or 0.1% BSA in PBS (gray) for 30 minutes, followed by incubation with secondary antibodies. BSA, bovine serum albumin; CHO, Chinese hamster ovary; PBS, phosphate-buffered saline.

Cross-reaction of PMab-231 to PDPNs of the other species using flow cytometry. CHO-K1 cells transfected with PDPNs of the other species were treated with PMab-231 (red line) or each positive control (blue line) at a concentration of 1 μg/mL or 0.1% BSA in PBS (gray) for 30 minutes, followed by incubation with secondary antibodies.
A kinetic analysis performed using flow cytometry assessed the interaction of PMab-231 with CHO/tigPDPN and CHO/cPDPN cells. KD of PMab-231 for CHO/tigPDPN and CHO/cPDPN cells were determined to be 1.2 × 10−8 and 1.9 × 10−8, respectively, indicating moderate affinity for CHO/tigPDPN and CHO/cPDPN cells.
Western blotting performed using PMab-231 demonstrated that PMab-231 detected broad bands of tigPDPN and cPDPN between 35 and 48 kDa in CHO/tigPDPN and CHO/cPDPN cells, respectively (Fig. 4). LpMab-17, an anti-LP tag mAb and PMab-1, an anti-MAP tag mAb, detected each band in CHO/tigPDPN and CHO/cPDPN cells, respectively.(24,32)

Western blotting. Cell lysates of CHO-K1, CHO/tigPDPN, and CHO/cPDPN (10 μg) electrophoresed and transferred onto PVDF membranes. The membranes incubated with l μg/mL PMab-231, 10 μg/mL anti-LP tag (LpMab-17), 1 μg/mL anti-MAP tag (PMab-1), or 1 μg/mL anti-β-actin, and subsequently, with peroxidase-conjugated anti-mouse or -rat IgG. cPDPN, cat PDPN; PVDF, polyvinylidene difluoride.
The immunohistochemical analyses revealed that PMab-231 stained the feline lungs (Fig. 5) and kidneys (Fig. 6), but PMab-231 did not react with lymphatic endothelial cells (data not shown). These results indicate that PMab-231 is useful for the detection of cPDPN using immunohistochemistry. Because tigPDPN and cPDPN shared 99% homology, PMab-231 may be useful for the detection of tigPDPN using immunohistochemistry.
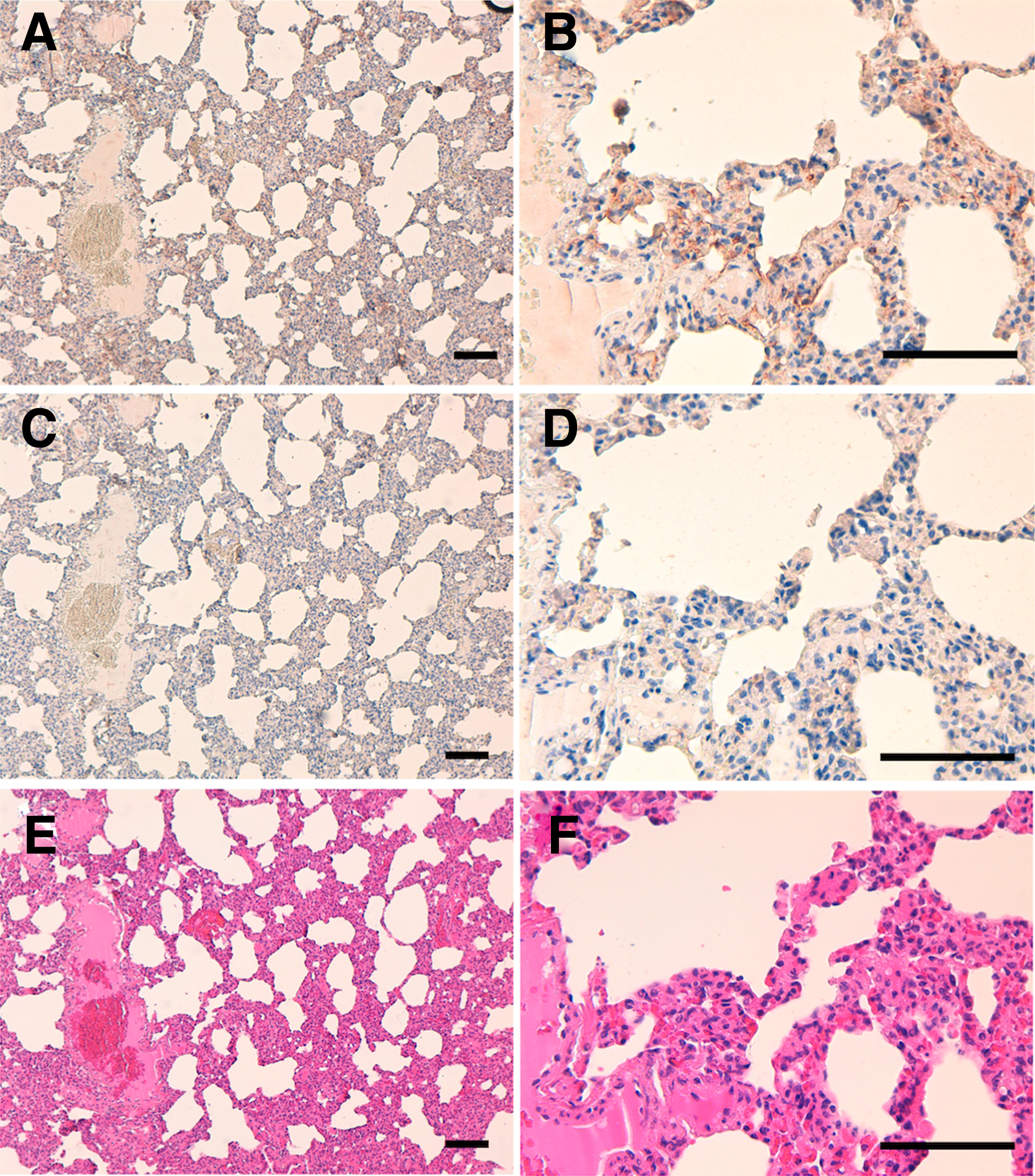
Immunohistochemical analyses against the feline lungs. Tissue sections of the feline lungs incubated with 5 μg/mL PMab-231

Immunohistochemical analyses against the feline kidneys. Tissue sections of the feline kidneys incubated with 5 μg/mL PMab-231
In conclusion, we established a mAb against tigPDPN, PMab-231, which is suitable for use in flow cytometry, western blotting, and immunohistochemical analyses. The epitope of PMab-231 is required to be further investigated to clarify the sensitivity and specificity of PMab-231 against tigPDPN. It is expected that PMab-231 would prove useful for the elucidation of the pathophysiological functions of tigPDPN in future studies.
Footnotes
Acknowledgments
We thank Miyuki Yanaka, Masato Sano, Kayo Hisamatsu, Saori Handa, and Yoshimi Nakamura for their excellent technical assistance. This research was supported in part by AMED under grant numbers JP18am0101078 (Y.K.), JP18am0301010 (Y.K.), and JP18ae0101028 (Y.K.), and by JSPS KAKENHI grant number 17K07299 (M.K.K.) and 16K10748 (Y.K.).
Author Disclosure Statement
Y.K. received research funding from Zenoaq Resource Co., Ltd. The other authors have no conflict of interest.