
Abstract
Diacylglycerol kinase (DGK) converts diacylglycerol (DG) into phosphatidic acid (PA). Both DG and PA serve as lipidic second messengers; therefore, DGK plays a critical role in regulating the balance of two signaling pathways mediated by DG and PA in cellular functions. DGK gamma (DGKγ), 1 of the 10 mammalian DGK isozymes, has been reported to be involved in membrane traffic, mast cell function, and leukemic cell differentiation. We previously developed a novel anti-DGKγ monoclonal antibody (mAb), DgMab-6, which is very useful in immunocytochemistry of human cultured cells. In this study, we characterized the binding epitope of DgMab-6 using Western blotting, and Glu12 is important for facilitating the DgMab-6 binding to the DGKγ protein. These results might lead to further development for sensitive and specific mAbs against DGKγ.
Introduction
Diacylglycerol kinase (DGK) phosphorylates diacylglycerol (DG) to produce phosphatidic acid (PA).(1,2) DG is a neutral lipid derived from various sources, including phosphatidylinositol 4,5-bisphosphate and phosphatidylcholine, and it serves as a second messenger that activates the conventional and novel types of the protein kinase C (PKC) family, RasGRP, Unc-13, and canonical transient receptor potential channels.(2,3) PA functions as a messenger molecule that activates the hypoxia-inducible factor-1α, atypical PKCζ, and mammalian target of rapamycin. DGK constitutes an enzyme family comprising 10 isozymes of the mammalian species.(1,2) Each isozyme has a distinct molecular structure and subcellular localization pattern. DGK gamma (DGKγ) was first isolated in rat(4) and human,(5) and is classified into type I DGK, which contains two EF hand motifs (Ca2+-biding site), two Zn fingers (DG-binding C1 domain), and a catalytic domain. DGKγ has been reported to be involved in a variety of cell functions including antigen-induced mast cell degranulation,(6) leukemic cell differentiation,(7) and membrane traffic between the endoplasmic reticulum and Golgi apparatus.(8)
We previously reported a novel antihuman DGKγ monoclonal antibody (mAb), DgMab-6, which is very useful in immunocytochemical analysis of cultured cells. In this study, we characterize the binding epitope of DgMab-6 using Western blotting.
Materials and Methods
Plasmid of DGKγ
Human DGKγ cDNA was synthesized, and was subcloned into an expression vector, pMAL-c2 (New England Biolabs, Inc., Beverly, MA), with PA tag (GVAMPGAEDDVV)(9) using the In-Fusion HD Cloning Kit (Takara Bio, Inc., Shiga, Japan); the resultant construct was named pMAL-c2-DGKγ-PA. The deletion mutants of DGKγ produced using PCR were subcloned into pMAL-c2 with PA tag using the In-Fusion HD Cloning Kit. The substitution of DGKγ amino acids (aas) 6–20 with alanine in dC200 of DGKγ was performed using the QuikChange Lightning Site-Directed Mutagenesis Kit (Agilent Technologies, Inc., Santa Clara, CA). These constructs were verified using direct DNA sequencing.
Western blotting
Competent Escherichia coli TOP-10 cells (Thermo Fisher Scientific, Inc., Waltham, MA) were transformed and cultured overnight at 37°C in Luria–Bertani medium (Thermo Fisher Scientific, Inc.) containing 100 μg/mL ampicillin (FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan). The cell pellets were resuspended in phosphate-buffered saline containing 1% Triton X-100 and 50 μg/mL aprotinin (Sigma-Aldrich Corp., St. Louis, MO). Lysates were boiled in sodium dodecyl sulfate (SDS) sample buffer (Nacalai Tesque, Inc., Kyoto, Japan). About dC80 lysates were immunoprecipitated using amylose resin (New England Biolabs, Inc.) and boiled in SDS sample buffer (Nacalai Tesque, Inc.). The samples were electrophoresed on 5%–20% polyacrylamide gels (Nacalai Tesque, Inc.) and transferred onto a polyvinylidene difluoride (PVDF) membrane (Merck KGaA, Darmstadt, Germany). After blocking with 4% skim milk (Nacalai Tesque, Inc.) for 1 hour, the membrane was incubated with DgMab-6 for 1 hour, followed by peroxidase-conjugated antimouse IgG (1:2000 dilution; Agilent Technologies, Inc.) for 1 hour. The membrane was also incubated with NZ-1 (anti-PA tag)(10) for 1 hour, followed by biotin-conjugated antirat IgG (1:1000 dilution; Agilent Technologies, Inc.) for 30 minutes, and further incubated with VECTASTAIN Elite ABC HRP Kit (Vector Laboratories, Inc., Burlingame, CA) for 30 minutes. The membrane was finally developed with the ImmunoStar LD Chemiluminescence Reagent (FUJIFILM Wako Pure Chemical Corporation) using the Sayaca-Imager (DRC Co., Ltd., Tokyo, Japan). All procedures of Western blotting were performed at room temperature.
Results
We herein characterized the binding epitope of DgMab-6 using Western blotting. As shown in Figure 1, we produced 19 C-terminal deletion mutants of DGKγ (i.e., dC380, dC360, dC340, dN320, dC300, dC280, dC260, dC240, dC220, dC200, dC180, dC160, dC140, dC120, dC100, dC80, dC60, dC40, and dC20). Western blotting demonstrated that DgMab-6 detected all deletion mutants, which were also detected by an anti-PA tag mAb, NZ-1 (Fig. 1A), indicating that the N-terminus of the DgMab-6 epitope exists between 1st aa and 20th aa.
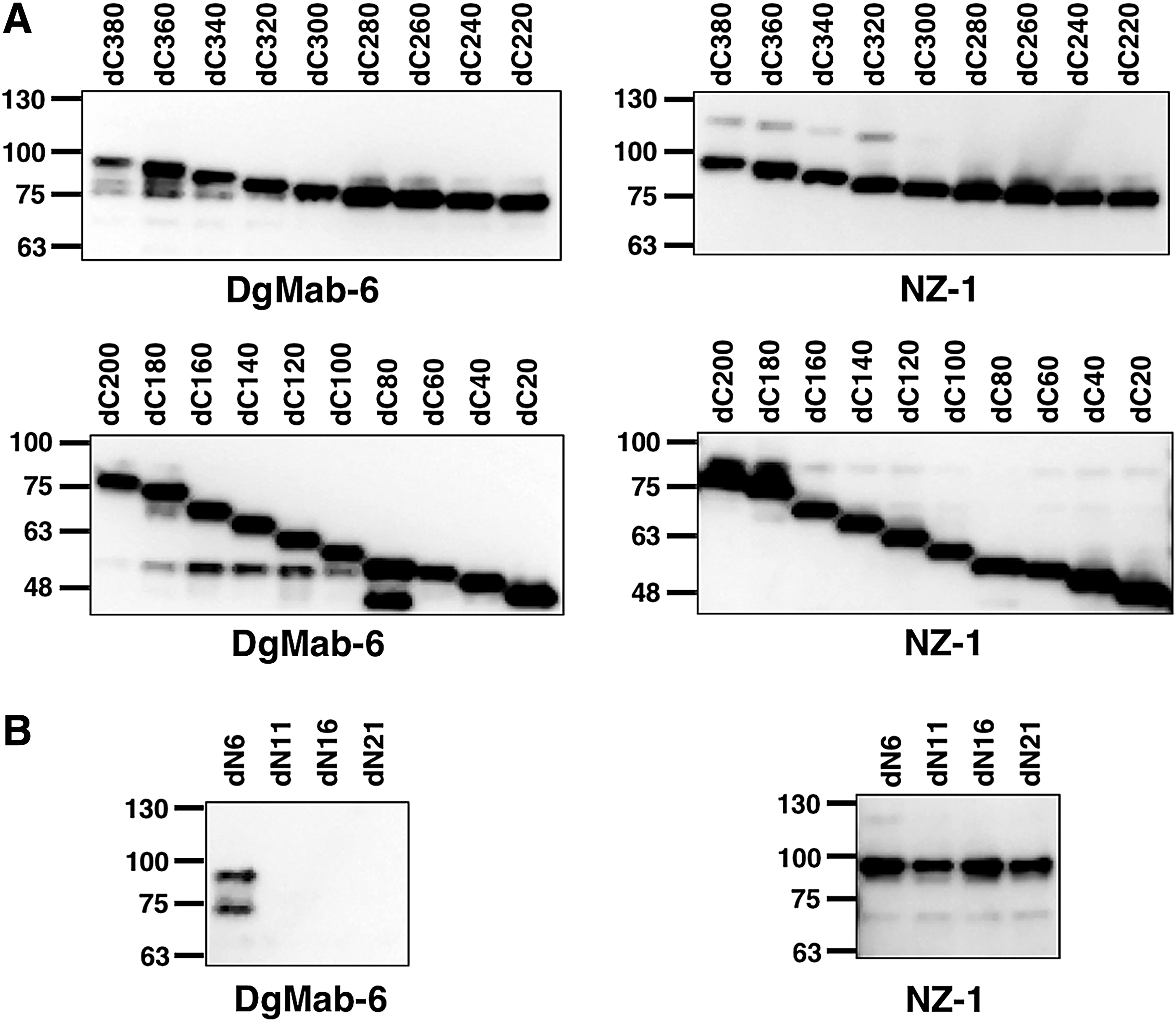
Production of DGKγ deletion mutants.
Western blotting of the additional deletion mutants of DGKγ—that is, dN6, dN11, dN16, and dN21—demonstrated that DgMab-6 detected dN6, but not dN11, dN16, and dN21, although all deletion mutants were detected by an anti-PA tag mAb, NZ-1 (Fig. 1B), thereby indicating that the N-terminus of the DgMab-6-epitope exists between 6th aa and 20th aa of DGKγ (Fig. 1B). These results are summarized in Figure 2.

Schematic illustration of DgMab-6 epitope. Black bars, deletion mutants detected by DgMab-6. hDGKγ, human DGKγ; MBP, maltose-binding protein.
Accordingly, we produced the following 15 DGKγ point mutants using human DGKγ-dC200: W6A, V7A, S8A, L9A, T10A, P11A, E12A, E13A, F14A, D15A, Q16A, L17A, Q18A, K19A, and Y20A. Western blotting demonstrated that the anti-PA tag mAb, NZ-1, detected all point mutants (Fig. 3A). In contrast, DgMab-6 did not detect E12A. These results are summarized in Figure 3B.
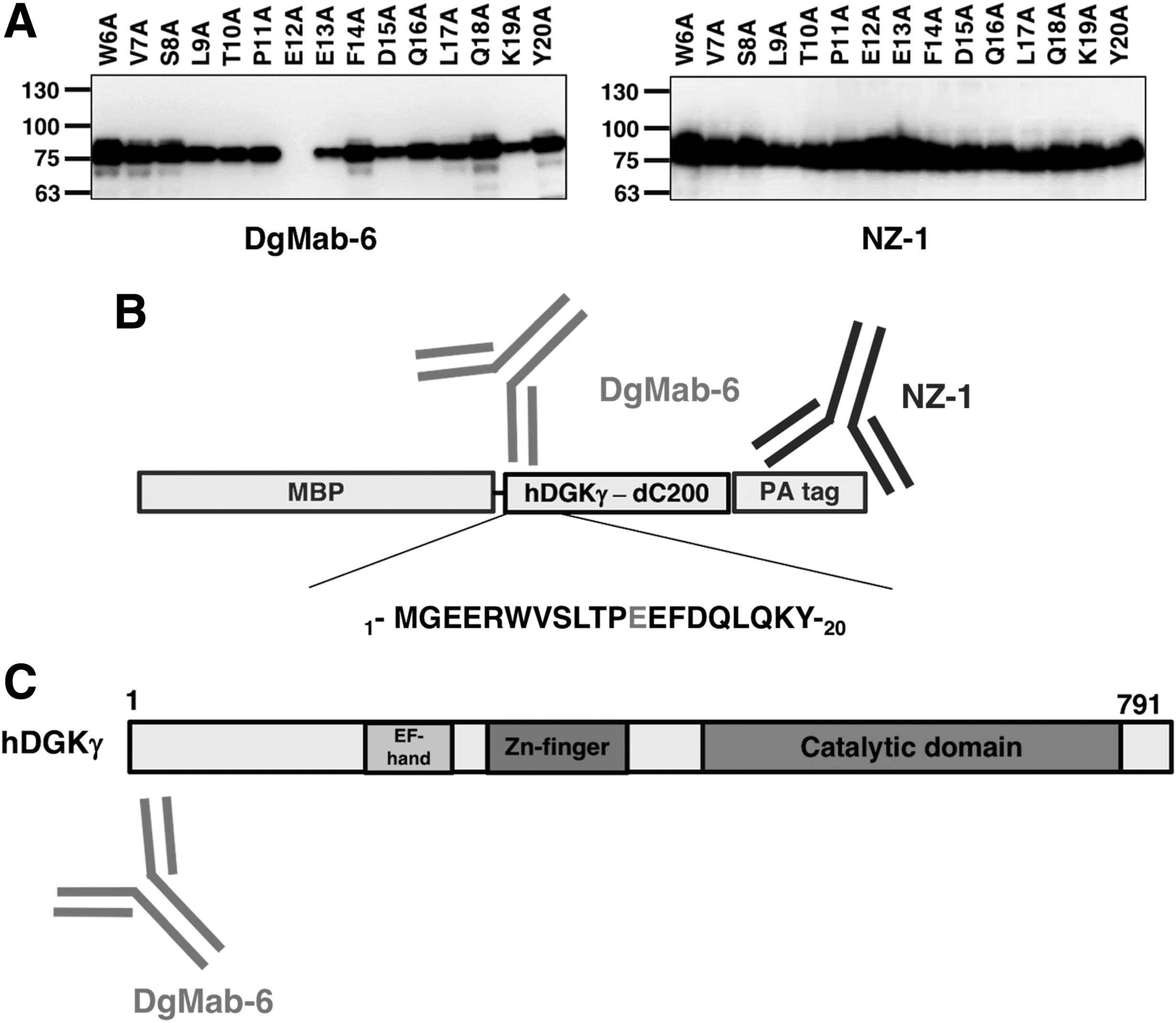
Epitope mapping of DgMab-6 using point mutants of DGKγ.
Discussion
Immunohistochemical examination using anti-DGKγ antibodies on rat tissues demonstrated cytoplasmic localization of DGKγ in several neurons of the brain.(4,11) In contrast, morphological examination on human cells or tissues has not yet been performed because highly specific and sensitive mAbs against human DGKγ had not been established. Recently, the recombinant MBP-DGKγ-PA was captured using NZ-1-Sepharose and efficiently eluted with PA tag peptide.(9) Mice were then immunized with MBP-DGKγ-PA to develop novel anti-DGKγ mAbs. Using ELISA, the culture supernatants of hybridomas were screened for binding to recombinant MBP-DGKγ-PA. Finally, DgMab-6 (mouse IgG1, kappa) was established. Immunocytochemical analysis revealed that immunoreactivity for DgMab-6 is detected as granular pattern in the cytoplasm in HL-60 cells, indicating that a novel anti-DGKγ mAb, DgMab-6, sensitively and specifically reacted with human DGKγ using immunocytochemical analysis. Previously, we further developed DaMab-2 (mouse IgG1, kappa), a specific mAb against DGKα, which is extremely useful in immunocytochemical analysis.(12) The binding epitopes of DaMab-2 were determined to be Cys246, Lys249, Pro252, and Cys253 of DGKα using Western blot analysis.(13) Because many cysteine residues are included in DGKα (24 cysteins in 735 aas; 3.3%), we could not synthesize DGKα peptides, which easily aggregate. Of interest, DaMab-2 epitope was included in Zn finger, and two cysteine residues such as Cys246 and Cys253 are involved in the DaMab-2 epitope. In this study, using Western blotting, we also characterized the binding epitope of DgMab-6 for DGKγ. DgMab-6 did not detect E12A mutant (Fig. 3A), indicating that Glu12 is critical for DgMab-6 binding to DGKγ (Fig. 3B). This epitope exists in the N-terminal domain of DGKγ (Fig. 3C). These findings could be applied for the production of more functional anti-DGKγ mAbs.
Footnotes
Acknowledgments
This research was supported, in part, by AMED under Grant numbers JP18am0101te078 (Y.K.), JP18am0301010 (Y.K.), and JP18ae0101028 (Y.K.), and by JSPS KAKENHI Grant number 17K07299 (M.K.K.) and Grant number 16K10748 (Y.K.).
Author Disclosure Statement
Y.K. received research funding from Ono Pharmaceutical Co., Ltd. The other authors have no conflict of interest.