
Abstract
Podoplanin (PDPN) is a mucin-type membrane glycoprotein, and possesses three platelet aggregation-stimulating (PLAG) domains: PLAG1, PLAG2, and PLAG3 at the N-terminus of PDPN, and one or two PLAG-like domains (PLDs) in the middle of PDPN. PDPN is expressed on normal tissues, such as podocytes of the kidney and type I alveolar cells of the lung, and is also overexpressed in numerous malignant cancers. Previously, we reported a novel anti-alpaca podoplanin (aPDPN) monoclonal antibody (mAb), PMab-225, using Cell-Based Immunization and Screening (CBIS) method. PMab-225 specifically detected aPDPN-overexpressed Chinese hamster ovary (CHO)-K1 (CHO/aPDPN) cells using flow cytometry and western blotting, and strongly stained alpaca tissues such as lung type I alveolar cells by immunohistochemistry. However, the specific binding epitope of aPDPN for PMab-225 remains unclear. Thus, in this study, a series of deletion or point mutations of aPDPN were utilized for investigating the binding epitope of PMab-225 using flow cytometry. The analysis of deletion mutants showed that N-terminus of PMab-225 epitope might exist between 80 amino acid (aa) and 85 aa of aPDPN. Furthermore, the analysis of point mutants demonstrated that Thr84 of aPDPN, which exists in PLD, could be included in the critical epitope of PMab-225.
Introduction
Podoplanin (PDPN/T1alpha/Aggrus), a type I transmembrane glycoprotein, is expressed in normal tissues, such as lymphatic endothelial cells, type I lung alveolar cells, and renal podocytes.(1–3) PDPN has been also known as a platelet aggregation-inducing factor in highly metastatic tumor cells.(1,4) PDPN is frequently overexpressed in various malignant tumors, including several squamous cell carcinomas, mesothelioma, glioblastoma, and osteosarcoma.(5–7) PDPN possesses three platelet aggregation-stimulating (PLAG) domains: PLAG1, PLAG2, and PLAG3; these domains exist at the N-terminus of PDPN.(8) Furthermore, one or two PLAG-like domains (PLDs) have been reported to be present in the middle of PDPN.(9) PLAG1–3 and/or PLD elicit tumor cell-induced platelet aggregation response through the activation of C-type lectin-like receptor 2 (CLEC-2) to facilitate cancer progression and metastasis.(1,3) These results imply the importance of the developing anti-PDPN monoclonal antibodies (mAbs) for cancer therapeutic treatment.
Alpaca (lama pacos) is a livestock that is used in various applications in Latin America. This animal has also been used for the production of antigen-specific single domain antibodies (nanobodies).(10,11) However, membrane proteins of alpaca have not been investigated due to the lack of specific antibodies. Because PDPN is important for blood/lymphatic vessel separation,(12,13) high endothelial venule integrity,(14) and embryonic cardiac development,(15) sensitive and specific mAbs against alpaca PDPN (aPDPN) are necessary to analyze the expression and function of aPDPN. Recently, we developed a novel anti-aPDPN mAb, PMab-225, using Cell-Based Immunization and Screening (CBIS) method. PMab-225 specifically detected aPDPN by flow cytometry, western blotting, and immunohistochemical analysis.(16) However, the binding epitope of PMab-225 for aPDPN remains to be clarified. Thus, this study aimed to identify the epitope of PMab-225 using deletion mutants or point mutants of aPDPN.
Materials and Methods
Cell lines
Chinese hamster ovary (CHO)-K1 was obtained from the American Type Culture Collection (ATCC, Manassas, VA). The aPDPN mutation plasmids containing N-terminal RAP16 tag were transfected into CHO-K1 cells using Lipofectamine LTX (Thermo Fisher Scientific, Inc., Waltham, MA). The RAP16 tag comprises 16 amino acids (aa; GPGDDMVNPGLEDRIE).(17) Transiently transfected cells with deletion/point-mutants were cultured in Roswell Park Memorial Institute (RPMI) 1640 medium (Nacalai Tesque, Inc., Kyoto, Japan), supplemented with 10% heat-inactivated fetal bovine serum (Thermo Fisher Scientific, Inc.), 100 units/mL of penicillin, 100 μg/mL of streptomycin, and 25 μg/mL of amphotericin B (Nacalai Tesque, Inc.) at 37°C in a humidified atmosphere of 5% CO2 and 95% air.
Production of aPDPN mutants
The coding sequence of aPDPN was subcloned into a pCAG vector (FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan), and a RAP16 tag was added at the N-terminus. Deletion mutants of aPDPN sequence were performed using a HotStar HiFidelity PCR (polymerase chain reaction; Qiagen, Inc., Hilden, Germany) with oligonucleotides containing the desired mutations. Substitutions of amino acids to alanine in aPDPN sequence were conducted by QuikChange lightning site-directed mutagenesis kit (Agilent Technologies, Inc., Santa Clara, CA). PCR fragments bearing the desired mutations were inserted into pCAG vector using In-Fusion PCR cloning kit (Takara Bio, Inc., Shiga, Japan).
Flow cytometry
Transiently transfected CHO-K1 cells were detached by 0.25% trypsin/1 mM ethylenediaminetetraacetic acid (EDTA; Nacalai Tesque, Inc.) and collected in RPMI 1640 medium. After washing with 0.1% bovine serum albumin/phosphate buffered saline, the cells were incubated with anti-aPDPN antibody (PMab-225; 1 μg/mL) or control anti-RAP16 tag antibody (PMab-2; 1 μg/mL) for 30 minutes at 4°C. Alexa Fluor 488-conjugated anti-mouse IgG (1:1000; Cell Signaling Technology, Inc., Danvers, MA) were added to each cell, and incubated for 30 minutes at 4°C. Fluorescence data were collected using a Cell Analyzer EC800 (Sony Corp., Tokyo, Japan), and were analyzed using FlowJo (BD Biosciences, Franklin Lakes, NJ).
Results
Ten deletion mutants and wild type (WT) of aPDPN were generated (Fig. 1A). Transient transfectants were produced using CHO-K1 cells, including WT (corresponding to 27–162 aa), dN30 (corresponding to 30–162 aa), dN40 (corresponding to 40–162 aa), dN50 (corresponding to 50–162 aa), dN60 (corresponding to 60–162 aa), dN70 (corresponding to 70–162 aa), dN80 (corresponding to 80–162 aa), dN85 (corresponding to 85–162 aa), dN90 (corresponding to 90–162 aa), dN95 (corresponding to 95–162 aa), and dN100 (corresponding to 100–162 aa). All deletion mutants and WT of aPDPN containing N-terminal RAP16 tag were analyzed using flow cytometry for the epitope mapping of PMab-225. PMab-2 (an anti-RAP16 tag mAb) detected all deletion mutants of aPDPN (Fig. 1B, left), indicating that the expression level of each deletion mutant is very high. In contrast, PMab-225 did not react with dN85, dN90, dN95, and dN100 (Fig. 1B, right), suggesting that N-terminus of PMab-225 epitope might exist between 80 aa and 85 aa of aPDPN.
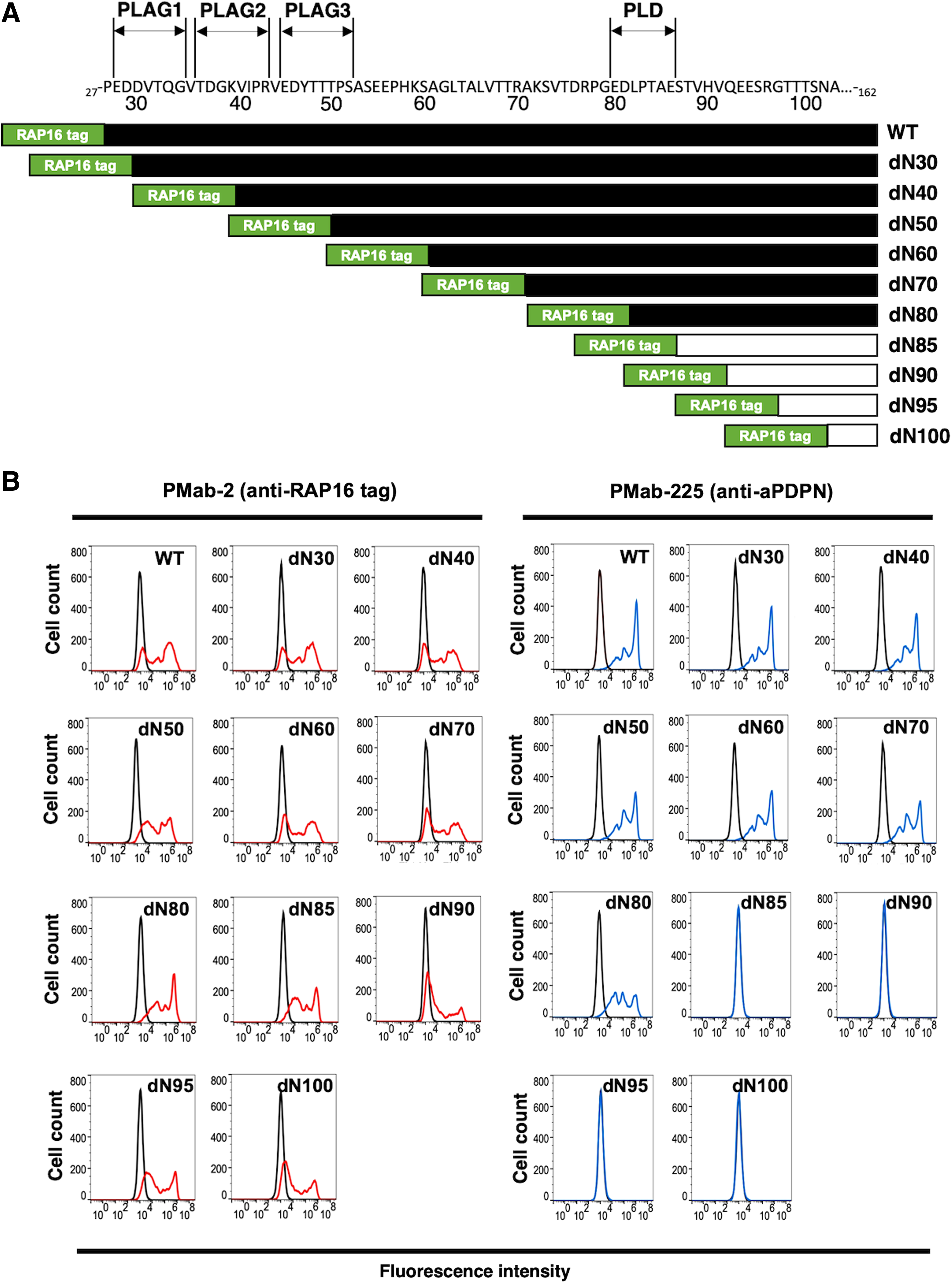
Epitope mapping of PMab-225 using deletion mutants of aPDPN.
Next, to identify the binding epitope of PMab-225, we produced a series of point mutations of aPDPN in CHO-K1 cells, including E80A, D81A, L82A, P83A, T84A, A85G, E86A, S87A, T88A, V89A, H90A, and V91A. PMab-2 reacted with all point mutants (Fig. 2A). In contrast, PMab-225 reacted with all these mutants, except for T84A (Fig. 2B), indicating that Thr84 of aPDPN could be included in the critical epitope of PMab-225.
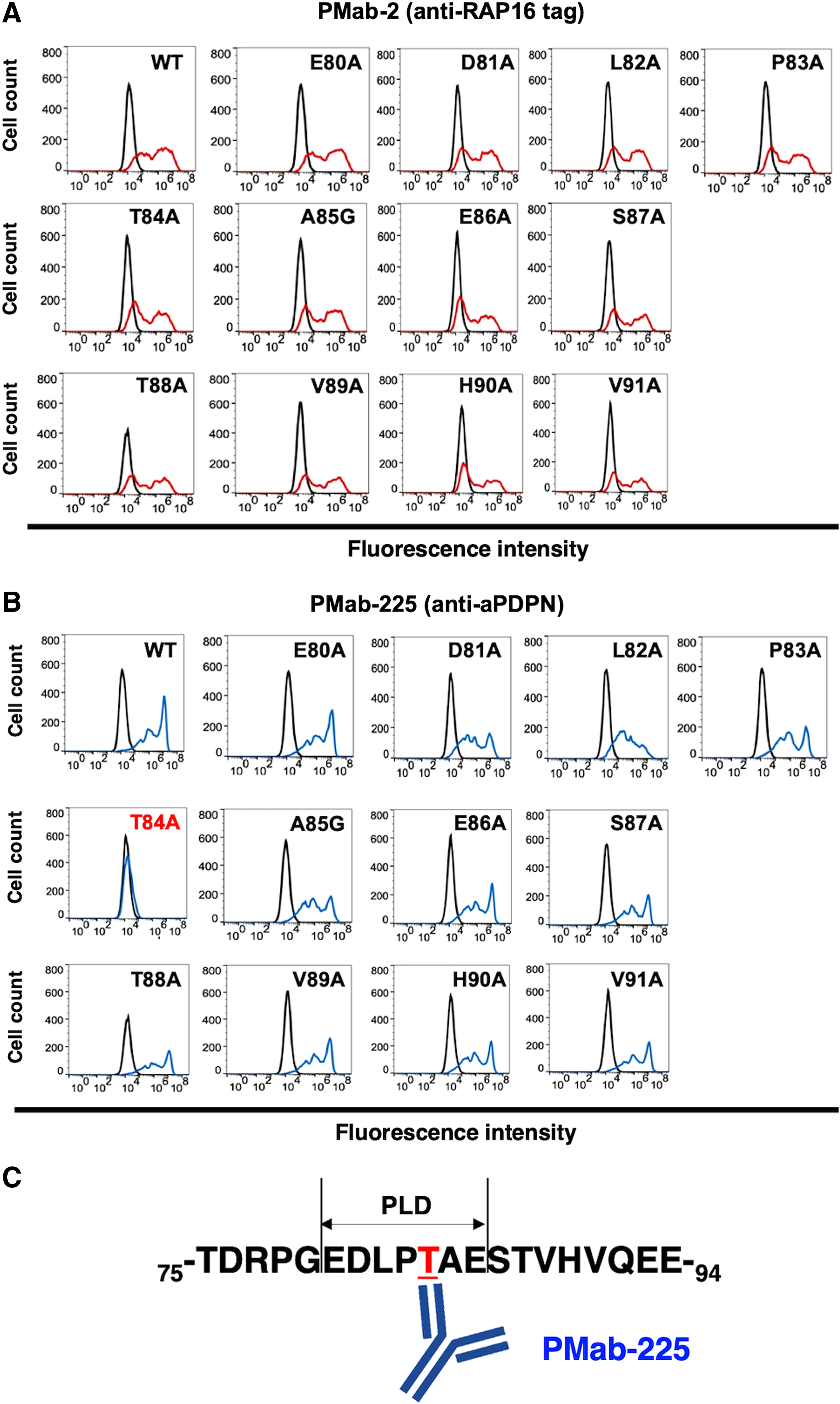
Epitope mapping of PMab-225 using point mutants of aPDPN.
Discussion
We have demonstrated that critical epitope of PMab-225 could be Thr84 of aPDPN using deletion mutants and point mutants of aPDPN. This result does not conclude that only Thr84 is a PMab-225-binding amino acid. In our experiences, the binding epitope of mAbs has been determined to be 5–13 aa(18,19); therefore, PMab-225 might bind to the peptide or the glycopeptide around Thr84 of aPDPN. We should perform X-ray crystallography to exactly determine the binding epitope of PMab-225 to aPDPN in the future study.
We have developed mAbs against many mammalian PDPNs, such as human,(20) mouse,(20) rat,(21) rabbit,(22) dog,(23) cat,(24) pig,(25,26) bovine,(27) horse,(28) Tasmanian devil,(29) tiger,(30) and bear(31) PDPNs. Almost all mAbs against PDPNs have been reported to react with the PLAG1–3 domains or PLD.(9,24,32) Actually, Thr84 of aPDPN exists in PLD (Fig. 2C). In the future study, we should investigate whether PMab-225 could neutralize the aPDPN-induced platelet aggregation.
In conclusion, Thr84 of aPDPN, which is located in PLD, is critical for PMab-225-specific binding to aPDPN. PMab-225 could be a useful tool for elucidating the pathophysiological function of aPDPN.
Footnotes
Author Disclosure Statement
The authors have no conflict of interest.
Funding Information
This research was supported in part by AMED under grant numbers JP19am0401013 (Y.K.), JP19am0101078 (Y.K.), and JP19ae0101028 (Y.K.), and by JSPS KAKENHI grant numbers 17K07299 (M.K.K.) and 19K07705 (Y.K.).