
Abstract
Understanding of the microenvironment of cancer plays a crucial role in cancer research. A tool is needed to evaluate the immune cells surrounding the cancer cells. This study establishes and evaluates a novel monoclonal antibody against canine CD8α (cCD8α). The antibody was produced by immunization of rats with cCD8α-expressing cells. After establishment and selection of hybridoma cells, the clone F3–B2 was established. The reactivity of F3–B2 was confirmed using cCD8α-overexpressing murine cells. Flow cytometric analysis also demonstrated that F3–B2 reacts with cCD8α naturally expressed in canine peripheral blood mononuclear cells and a canine T cell lymphoma cell line. The specimens of lymphoid tissue showed immunohistochemical staining for F3–B2. Moreover, we also found that F3–B2 exhibited reactivity against feline CD8. Thus, this antibody provides a good research tool to analyze CD8-positive cytotoxic lymphocytes in canine and feline tumors.
Introduction
Cancer is one of the main causes of death in older dogs. Some of the naturally occurring canine cancers have also received attention as an ideal model for human cancer. For instance, soft tissue sarcomas in dogs and humans have shown to share similar genetic complexity.(1) In addition, dogs with osteosarcoma show similar biological behavior and response to treatment to humans.(2) Canine lymphoma also shares many characteristics, such as incidence, biological behavior, and genetic aberrations, with human lymphoma.(3) The value of dogs as a lymphoma model is also supported by previous study, in which dogs with spontaneous B cell non-Hodgkin lymphoma were enrolled in a clinical trial of Bruton tyrosine kinase (Btk) inhibitor.(4)
The evaluation of cancer immunotherapy in dogs offers particular advantages in some kind of tumor, because immunocompetent dogs have an immunosuppressive cancer microenvironment that enables evaluation of the interaction between cancer and immune cells. Several studies have demonstrated that, as with human cancers, the immunosuppressive cancer microenvironment, such as regulatory T cells, tumor-associated macrophages, and myeloid-derived suppressor cells, exists and correlates with poor prognosis in canine patients with B cell lymphoma and mammary tumor.(5–7) Therefore, tools for evaluating canine immune cells are important. Several antibodies have been reported; however, antibodies which can detect canine CD8 via immunohistochemistry using formalin-fixed and paraffin-embedded (FFPE) samples are limited.(8–12) This study established a novel monoclonal antibody against canine CD8α (cCD8α), evaluated by flow cytometry and immunohistochemistry.
Materials and Methods
Cells
Rat kidney cell line, normal rat kidney epithelial cells (NPK), murine fibroblast cell line, NIH-3T3, retroviral packaging cell lines, PLAT-GP and PLAT-E, and feline T cell lymphoma cell line, 3201, were cultured in D10 complete medium (Dulbecco's modified Eagle's medium supplemented with high glucose, 10% fetal bovine serum (FBS), 100 U/mL penicillin, 100 μg/mL streptomycin, and 55 μM 2-mercaptoethanol). Canine T cell lymphoma cell line, UL-1, was cultured in R10 complete medium (RPMI1640 supplemented with 10% FBS, 100 U/mL penicillin, 100 μg/mL streptomycin, and 55 μM 2-mercaptoethanol). Peripheral blood mononuclear cells (PBMCs) were isolated from a healthy dog and were stimulated by interleukin (IL)-2 and phytohemagglutinin.
NRK cells stably expressing chimeric antigen receptor (CAR) were established via retrovirus transduction, as previously described.(13,14) A third-generation CAR construct was used in this study. The CAR construct was composed of the anti-CD20 single-chain variable fragment (scFv) (developed in our laboratory) linked to the cCD8α hinge and transmembrane domain, followed by the canine CD28 and 4–1BB costimulatory domain, and human CD3ζ signaling domain. For the coexpression of CARs together with fluorescent protein, the CAR construct was followed by a P2A peptide fused to Venus fragments. This CAR-P2A-Venus construct was ligated into pMx-IP retroviral vector. Retrovirus particles were generated by transient transfection of Plat-GP cells with the CAR-encoding retrovirus vector. Supernatants containing the retrovirus were collected after 48 hours, and used to transduce the NRK cells (NRK/CAR).
To obtain the cell line expressing the full length-cCD8α, the cCD8α gene was subcloned into a pMx-IP retroviral vector containing a puromycin resistance gene. Retrovirus particles were generated by transient transfection of PLAT-E cells with the cCD8α-encoding pMx-IP vector, and NIH-3T3 cells were transduced with supernatants containing cCD8α-expressing retrovirus, followed by puromycin selection (NIH-3T3/cCD8α). In parallel, NIH-3T3 cells were transduced with a retrovirus encoding the empty vector, as a control (NIH-3T3/emp).
CD8 knockout 3201 cells were generated using the CRISPR-Cas9 technology. Guide sequences targeting feline CD8 (5′-CACCGGTTCCGCTTATCGCCCGTGA-3′ and 5′-AAACTCACGGGCGATAAGCGGAACC-3′) were cloned into a lentiCRISPRv2 plasmid (gift from Feng Zhang [Addgene Cambridge, MA, plasmid #52961; 52961; RRID:Addgene_52961]). CRISPR plasmid was cotransfected with packaging plasmids, p8.9QV and pCVSVG, into HEK293T cells. After transfection to HEK293T cells, supernatants, including CRISPR lentivirus particles, were harvested, and 3201 cells were infected with the concentrated viral supernatant and cultured in the presence of puromycin (1 μg/mL) to select the stably transduced cells.
All cells were cultured in a humidified incubator at 37°C and 5% CO2.
Production of a rat monoclonal antibody
All animal studies were carried out in accordance with the Yamaguchi University Animal Care and Use Committee Regulations. A panel of antibodies against cCD8α was obtained by immunization with NRK/CAR cells, as previously described.(15) In brief, 1 × 107 NRK/CAR cells in 500 μL of phosphate-buffered saline (PBS) were emulsified with an equal amount of TiterMax Gold (CytRx, California), and injected intracutaneously into the hind footpads of 6-week-old Sprague-Dawley rats (SLC Japan, Shizuoka, Japan). Eight weeks after injection, cells were collected from popliteal lymph nodes and fused with P3U1 myeloma cells. Hybridoma cells that produced antibodies positive for cCD8α were identified by cell enzyme-linked immunosorbent assay and flow cytometry, and the resulting hybridoma cells were cloned by limiting dilution. After replacement of the culture media by serum-free media, the monoclonal antibody was purified using Protein G Sepharose 4 Fast Flow (GE Healthcare Japan, Tokyo, Japan).
Flow cytometry
Cells were collected and resuspended in fluorescence-activated cell sorting (FACS) buffer (PBS containing 2% FBS and 0.1% NaN3). A total of 2 × 105 cells were stained with each antibody for 30 minutes on ice. Cell surface expression of CD8α was detected by incubation with the established antibody, followed by incubation with DyLight 649-labeled anti-rat IgG antibody (BioLegend, San Diego, CA). In addition to the established antibody, the following antibodies were used: rat anti-dog CD8α APC (clone YCATE55.9; eBioscience, Inc., Vienna, Austria), rat anti-dog CD8α FITC (clone YCATE55.9; Bio-Rad Laboratories, Inc., Hercules, CA), and corresponding isotype control antibodies. For isotyping analysis of established antibody, cCD8α-expressing cells were stained with established antibody, followed by biotin-labeled anti-rat Ig antibodies specific to each subclass (BioLegend), and detected using Dylight 649-labeled streptavidin (BioLegend). Samples were analyzed using an Accuri C6 (BD Biosciences, San Diego, CA), and results were analyzed using FlowJo software (BD Biosciences).
Immunohistochemistry
FFPE sections from healthy dogs and cats were used for immunohistochemical staining, as described in our previous report.(16) After dewaxing, antigen retrieval proceeded in citrate buffer (pH 6.0) at 125°C for 30 seconds. In the case of feline sections, antigen retrieval was performed in citrate buffer (pH 9.0) at 125°C for 10 minutes. Then the sections were washed in distilled water and rinsed with PBS. Endogenous peroxidase activity was blocked with 3% hydrogen peroxide in methanol for 30 minutes at room temperature (RT). The sections were washed with distilled water, then rinsed with PBS. To block nonspecific protein activity, the sections were incubated for 30 minutes at RT with 5% skimmed milk and 5% bovine serum albumin (BSA) in PBS. After blocking nonspecific protein activity, the sections were rinsed with PBS, followed by incubation with primary antibody overnight at 4°C. All primary antibodies were diluted with 2% BSA in PBS. Rat anti-human CD3 antibody (clone CD3–12: Serotec, Oxford, United Kingdom), rabbit anti-human Granzyme B (Spring Bioscience Corp, Pleasanton, CA), the established antibody, and corresponding isotype control antibodies were used as primary antibody. Anti-human CD3 and anti-human Granzyme B antibodies were confirmed to cross-react with canine tissues in previous reports.(17,18) The antibody-treated sections were rinsed with PBS, followed by treatment with horseradish peroxidase-conjugated secondary antibodies (goat anti-rat IgG antibody for CD3 and the established antibody; goat anti-rabbit IgG antibody for Granzyme B; Histofine Simple Stain MAX PO: Nichirei Corporation, Tokyo, Japan) for 30 minutes at RT. The sections were rinsed with PBS, and color was developed with the peroxidase stain 3′,3′-diaminobenzidine (DAB Kit & Enhancer; Nacalai Tesque, Kyoto, Japan) for 5 minutes at RT. After the sections were washed in distilled water, they were counterstained for 5 minutes with Mayer's hematoxylin solution (Merck KGaA, Darmstadt, Germany), washed in distilled water, dehydrated in graded baths of ethanol, cleared in xylene, and mounted with a xylene-based mounting solution (Matsunami, Osaka, Japan).
Results and Discussion
To establish the monoclonal antibody against cCD8α, we immunized rats with cCD8α-expressing NRK cells. In our laboratory, to develop a novel cancer therapy for canine cancer, an immune cell therapy using CAR-expressing T (CAR-T) cells was previously developed (in submission). CAR-T cells were developed by transducing the CAR construct to have the antibody scFv and cCD8α as extracellular domains (Fig. 1A), and this CD8α was considered useful for immunization to induce antibodies against CD8α. CAR-expressing cells were used for immunization of rats, because the original purpose of the study was to establish an antibody that could detect CAR. Since the CAR construct used has scFv and CD8α as an extracellular domain, there was a possibility that antibodies against these two molecules were produced. Therefore, in this study, we focused on the antibody against cCD8α. Also, after hybridoma selection and cloning, a single hybridoma clone (F3–B2) was established.
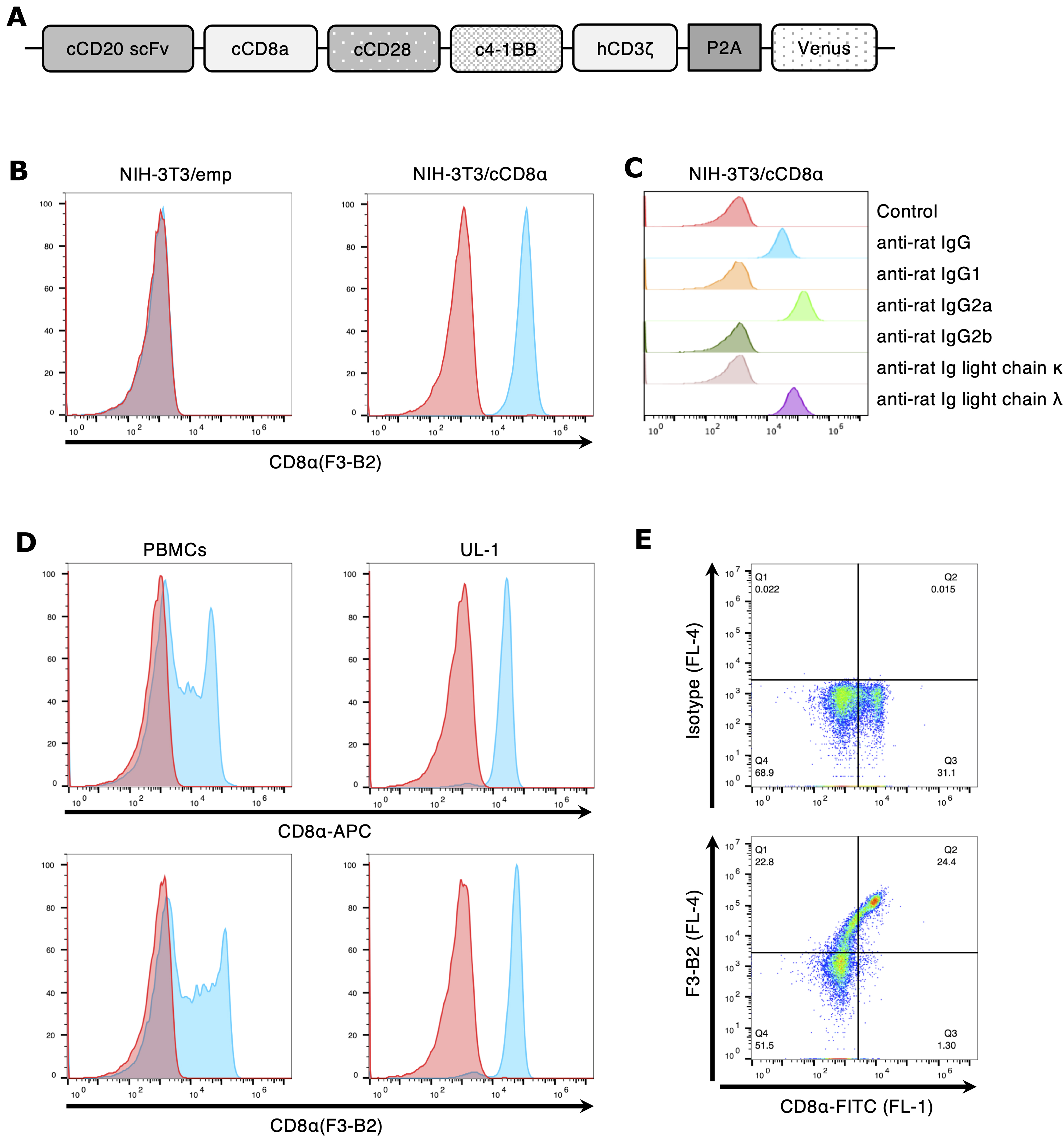
Generation and characterization of anti-cCD8α antibody.
First, the NIH-3T3/emp and NIH-3T3/cCD8α cells were stained with F3–B2 and isotype control antibodies, and it was confirmed that F3–B2 detected only cCD8α-expressing NIH-3T3 cells, whereas F3–B2 did not detect NIH-3T3/emp (Fig. 1B). Next, antibody isotyping of F3–B2 was performed using flow cytometry, and the subclass of F3–B2 was determined to be IgG2a lambda (Fig. 1C). To determine whether this antibody can detect endogenous CD8α expressed on canine cells, PBMCs and the canine T cell lymphoma cell line UL-1 were stained with F3–B2 antibody. As shown in Figure 1D, F3–B2 reacted with both PBMCs and UL-1 cells (bottom panels). Also, F3–B2 and the commercially available anti-cCD8α antibody (CD8α-APC) had almost the same reactivity (top panels). Double staining analysis by flow cytometry revealed that cells of the same population of canine PBMCs were stained positively with both F3–B2 and commercially available anti-cCD8α antibody (CD8α-FITC) (Fig. 1E). These results indicate that the established monoclonal antibody, clone F3–B2, recognizes cCD8α. In addition, F3–B2 showed almost the same reactivity as commercially available anti-cCD8α antibody (Fig. 1E). However, the double staining analysis showed that there was a cell population that became stained with both antibodies (Fig. 1E). This result indicated that the epitopes of these antibodies did not overlap.
Next, F3–B2 was assessed to determine whether it could be used for immunohistochemistry. The results of the immunohistochemical staining are shown in Figure 2. All lymphoid tissue specimens were positively stained by F3–B2 antibody. Many F3-B2-positive cells were detected in the lymph node and thymus, while only a few cells were positively stained in the spleen. The specimens were also positively stained for CD3 and Granzyme B. In lymph node specimens, staining patterns were similar, and mainly cells in the paracortical area were positive for CD3, Granzyme B, and CD8α (Fig. 2A). From these results, F3–B2 was also shown to be adaptable for immunohistochemistry using paraffin-embedded tissues.

Immunohistochemical analysis using established anti-cCD8α antibody. Tissue specimens from healthy dogs were stained immunohistochemically with anti-human CD3, anti-human Granzyme B, and anti-cCD8α (F3–B2). Representative immunostaining of lymph node
Finally, F3–B2 was also evaluated whether this antibody can detect feline CD8. As shown in Figure 3A, lymph node tissue from healthy cats showed positive staining for F3–B2. Feline CD8 knockout 3201 cells (3201/fCD8ko) were established to confirm that F3–B2 specifically recognize feline CD8. FACS analysis revealed that F3–B2 can react with wild-type 3201 cells, whereas F3–B2 cannot react with 3201/fCD8ko cells (Fig. 3B). These results suggest that F3–B2 can be used to detect feline CD8 in both immunohistochemical staining and flow cytometry.

F3–B2 also shows reactivity against feline CD8.
In this study, we established a novel antibody against cCD8α, and demonstrated that this antibody can be used in not only flow cytometry but also immunohistochemistry using FFPE sections. CD8 is the main marker of cytotoxic T lymphocytes, which play an important role in cancer regression. In addition, the significance of tumor-infiltrating lymphocytes as a prognostic factor has been reported in canine tumors.(19,20) Franzoni et al. evaluated tumor-infiltrated CD8+ T lymphocytes in canine mammary-gland tumors using immunohistochemistry.(6) However, they used freshly frozen samples because anti-CD8 antibodies that can be used in canine FFPE samples are limited. Antibody detectable in FFPE samples enables further analysis from archive samples.
In conclusion, we established the antibody against cCD8α and confirmed that this antibody can be used in immunohistochemistry using FFPE samples. The novel anti-cCD8α antibody of the present study, clone F3–B2, enables a more precise evaluation of canine CD8+ cells, providing a new method for determining the immune responses in dogs and cats.
Footnotes
Acknowledgments
We acknowledge the technical expertise of the DNA Core Facility of the Center for Gene Research, Yamaguchi University. We thank Enago for the English language review.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.