
Abstract
Diacylglycerol kinase (DGK) η is classified as a type II DGK and catalyzes diacylglycerol phosphorylation to produce phosphatidic acid. DGKη has been reported to be highly expressed in the hippocampus and cerebellum. Although a DGKη-specific monoclonal antibody (mAb) is necessary to reveal the association between the expression of DGKη and diseases, an anti-DGKη mAb for immunohistochemistry has not been developed. Recently, we established a specific antihuman DGKη (hDGKη) mAb, DhMab-1 (mouse IgG2a, kappa). For epitope mapping of DhMab-1, here we produced deletion or point mutants of hDGKη and performed Western blotting to determine the binding epitope of DhMab-1. DhMab-1 reacted with the dN755 mutant, but not with the dN760 mutant, indicating that the N-terminus of the DhMab-1 epitope is mainly located between amino acids 755 and 760 of the protein. A more detailed analysis using point mutants demonstrated that seven mutants, that is, A751G, I755A, D756A, P757A, D758A, L759A, and D760A, were not detected by DhMab-1. These results indicate that Ala751, Ile755, Asp756, Pro757, Asp758, Leu759, and Asp760 are important for DhMab-1 binding to hDGKη.
Introduction
Diacylglycerol kinase (DGK) plays a key role in the G-protein–mediated signaling pathway, which is an important signaling cascade in mammalian cells.(1,2) DGK is an enzyme that phosphorylates diacylglycerol to produce phosphatidic acid. In turn, phosphatidic acid functions as a second messenger that mediates intracellular Ca2+ levels and regulates the mTOR-mediated signaling pathway.(3,4) The DGK family consists of 10 isozymes in mammalian species, with each isozyme containing two to three C1 domains, while sharing a common catalytic domain.(1) The DGK family is further divided into five subtypes according to their subtype-specific functional domains.(5) DGKη is classified as a type II DGK. It contains a pleckstrin homology (PH) domain and two C1 domains, a catalytic domain, and an accessory domain.(6) The PH domain is a phosphatidylinositol-binding domain. In particular, the PH domain of DGKη binds to phosphatidylinositol 4,5-bisphosphate and regulates the subcellular localization of DGKη.(7)
Recently, we established a specific antihuman DGKη (hDGKη) monoclonal antibody (mAb), DhMab-1 (mouse IgG2a, kappa). In this study, we performed epitope mapping to identify the binding epitope of DhMab-1 using deletion and point mutations of hDGKη.
Materials and Methods
Plasmid preparation and production of recombinant DGKη protein
Synthesized DNA (Eurofins Genomics KK, Tokyo, Japan) encoding hDGKη (accession No. AB078967) plus a C-terminal PA tag (GVAMPGAEDDVV)(8,9) was subcloned into the expression vector pMAL-c2 (New England Biolabs, Inc., Beverly, MA) using the In-Fusion HD Cloning Kit (Takara Bio, Inc., Shiga, Japan). The PA tag is recognized by an anti-PA tag mAb (NZ-1).(10) The resulting construct was named pMAL-c2-hDGKη-PA. The deletion mutants of hDGKη were produced by polymerase chain reaction and were subcloned into pMAL-c2 with a PA tag using the In-Fusion HD Cloning Kit. The substitution of the amino acids of hDGKη with alanine at dN705 of hDGKη was performed using the QuikChange Lightning Site-Directed Mutagenesis Kit (Agilent Technologies, Inc., Santa Clara, CA). These constructs were verified by direct DNA sequencing.
Competent Escherichia coli TOP-10 cells (Thermo Fisher Scientific, Inc., Waltham, MA) were transformed with the pMAL-c2-hDGKη-PA plasmid. They were cultured overnight at 37°C in Luria-Bertani broth (Thermo Fisher Scientific, Inc.) containing 100 μg/mL ampicillin (Sigma-Aldrich Corp., St. Louis, MO). Cell pellets were resuspended in phosphate-buffered saline containing 1% Triton X-100 and 50 μg/mL aprotinin (Sigma-Aldrich Corp.). After sonication, the crude extracts were collected by centrifugation (9000 g, 30 minutes, 4°C). The lysates were passed through a 0.45 μm filter to remove any trace amounts of insoluble materials. Cleared lysates were mixed with NZ-1-Sepharose (1 mL bed volume) and incubated at 4°C for 2 hours under gentle agitation. The resin was then transferred to a column and washed with 20 mL Tris-buffered saline (pH 7.5). The bound protein was eluted with the PA tag peptide at room temperature in a stepwise manner (1 mL × 10 washes).
Hybridoma production
The Animal Care and Use Committee of Tohoku University approved all animal experiments. DhMab-1 was produced using the mouse medial iliac lymph node method. In brief, B6D2F1/Slc mice (Japan SLC, Inc., Shizuoka, Japan) were immunized by injecting 33 μg of the pMAL-c2-hDGKη-PA protein together with Freund's complete adjuvant (Sigma-Aldrich Corp.) into their footpad. Additional immunization with 60 μg of the pMAL-c2-hDGKη-PA protein was performed through the tail base. The lymphocytes were fused with mouse myeloma Sp2/0-Ag14 cells using polyethylene glycol. The culture supernatants were screened using enzyme-linked immunosorbent assay of the binding to the pMAL-c2-hDGKη-PA protein.
Western blot analyses
Lysates were boiled in sodium dodecyl sulfate sample buffer (Nacalai Tesque, Inc., Kyoto, Japan). The samples were electrophoresed on 5%–20% polyacrylamide gels (Nacalai Tesque, Inc.) and transferred onto a polyvinylidene difluoride membrane (Merck KGaA, Darmstadt, Germany). After blocking with 4% skim milk (Nacalai Tesque, Inc.) for 1 hour, the membrane was incubated with DhMab-1 (1 μg/mL) or NZ-1 (1 μg/mL) for 1 hour, followed by incubation with hydrogen peroxidase-conjugated antimouse immunoglobulins (1:2000 dilution; Agilent Technologies, Inc.) or HRP-conjugated antirat IgG (1:10,000 dilution; Sigma-Aldrich Corp.) for 1 hour. The membrane was developed with the ImmunoStar LD Chemiluminescence Reagent (FUJIFILM Wako Pure Chemical Corporation) using the Sayaca-Imager (DRC Co., Ltd., Tokyo, Japan). All Western blot procedures were performed at room temperature.
Results
Previously, we established DhMab-1 (mouse IgG2a, kappa), which can recognize hDGKη but not other related isozymes, such as DGKα, DGKγ, and DGKζ, in an enzyme-linked immunosorbent assay (data not shown). In this study, we performed epitope mapping to characterize the binding epitope of DhMab-1 using Western blot analysis.
We produced a C-terminal deletion mutant (dC583) and an N-terminal deletion mutant (dN584), as shown in Figure 1. First, we investigated whether DhMab-1 recognizes either dC583 or dN584 by Western blotting. The results of this experiment indicated that DhMab-1 interacts with dN584 (Fig. 2A).

Schematic illustration of the hDGKη deletion mutants. Black bars, deletion mutants detected by DhMab-1. DGKη, diacylglycerol kinase η; hDGKη, human DGKη; MBP, maltose-binding protein.
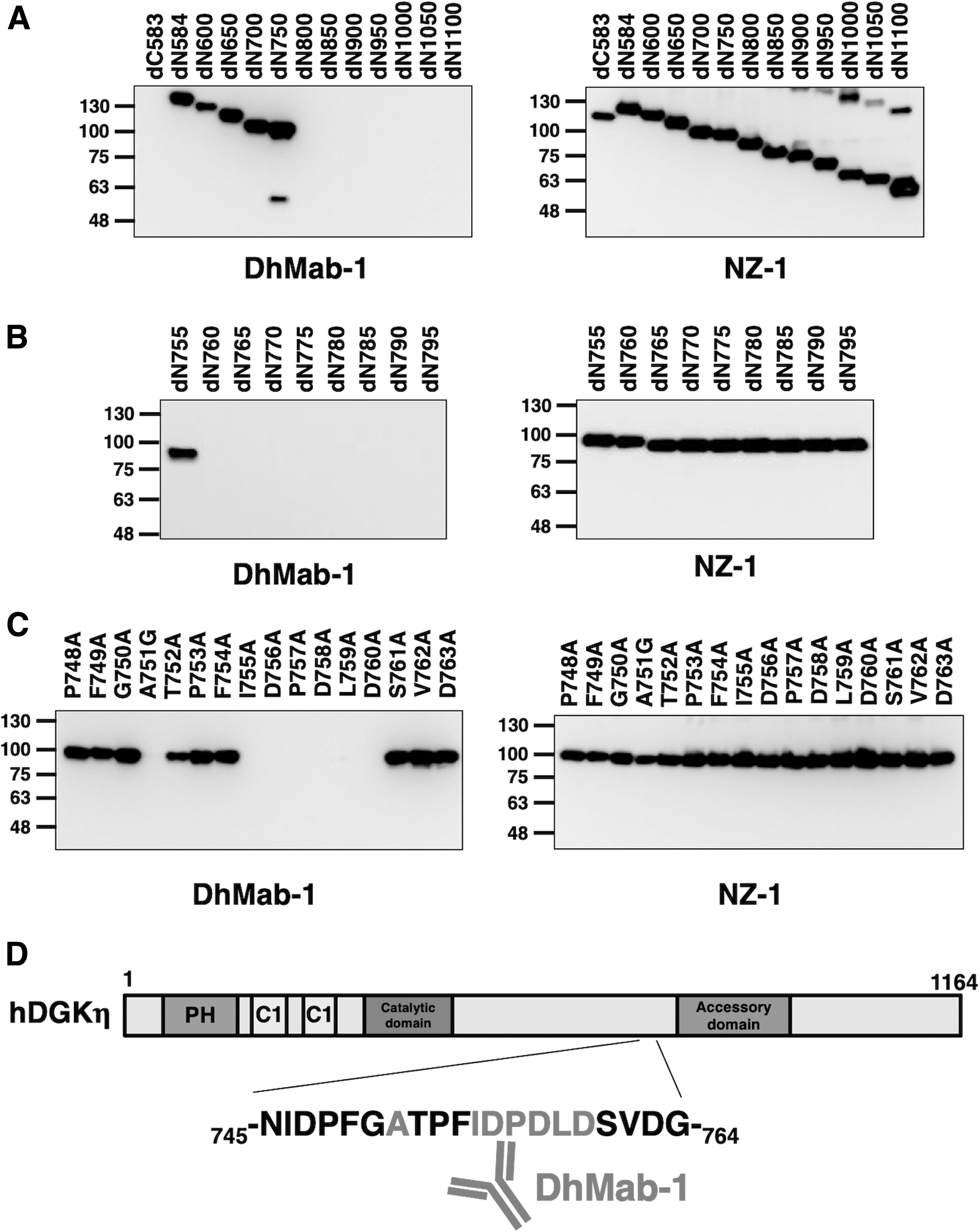
Epitope mapping of DhMab-1 using deletion mutants and point mutants of hDGKη.
We then produced an additional 11 N-terminal deletion mutants (dN600, dN650, dN700, dN750, dN800, dN850, dN900, dN950, dN1000, dN1050, and dN1100) of hDGKη (Fig. 1) and performed Western blotting. As shown in Figure 2A, DhMab-1 recognized dN600, dN650, dN700, and dN750, but not dN800, dN850, dN900, dN950, dN1000, dN1050, and dN1100. All of the deletion mutants were detected by the anti-PA tag mAb, NZ-1 (Fig. 2A). This result indicates that the DhMab-1 epitope is located between amino acids 750 and 800.
Next, we produced an additional nine N-terminal deletion mutants (dN755, dN760, dN765, dN770, dN775, dN780, dN785, dN790, and dN795) of hDGKη (Fig. 1). Western blotting demonstrated that DhMab-1 recognized dN755, but not dN760, dN765, dN770, dN775, dN780, dN785, dN790, and dN795, indicating that the DhMab-1 epitope is located between amino acids 755 and 760. All of the deletion mutants were detected by the anti-PA tag mAb, NZ-1 (Fig. 2B).
To investigate further the critical epitope of DhMab-1, we produced constructs representing 16 point mutants within hDGKη (P748A, F749A, G750A, A751G, T752A, P753A, F754A, I755A, D756A, P757A, D758A, L759A, D760A, S761A, V762A, and D763A). All hDGKη point mutants were recognized by NZ-1 (Fig. 2C). In contrast, DhMab-1 did not recognize A751G, I755A, D756A, P757A, D758A, L759A, and D760A (Fig. 2C), indicating that DhMab-1 binds to DGKη through the following seven amino acids: Ala751, Ile755, Asp756, Pro757, Asp758, Leu759, and Asp760. These results are summarized in Figure 2D.
Discussion
Previously, we established the anti-DGKα mAbs DaMab-2 and DaMab-8,(11,12) an anti-DGKγ mAb, DgMab-6,(13) and an anti-DGKζ mAb, DzMab-1.(14) We further determined their respective binding epitopes.(12,15–17) DaMab-2 and DaMab-8 bind to the Zn-finger domain and catalytic domain of DGKα, respectively.(12,15) DgMab-6 and DzMab-1 bind to the N-terminus of DGKγ and DGKζ, respectively.(16,17) Recently, we established a novel anti-hDGKη mAb, DhMab-1, which is useful for immunohistochemical analysis (data not shown). In this study, we identified the binding epitope of DhMab-1 by Western blotting. Seven amino acids of hDGKη, that is, Ala751, Ile755, Asp756, Pro757, Asp758, Leu759, and Asp760, are critical for DhMab-1 binding to the hDGKη protein. This epitope is located near the accessory domain of hDGKη (Fig. 2D).
DGKη is highly expressed in the hippocampus and cerebellum areas of the brain.(18) The DGKη mRNA is upregulated in patients with bipolar disorder.(19,20) DGKη is also highly expressed in lung cancer containing EGFR mutations, and knockdown of DGKη resulted in impaired growth of EGFR-mutant cell lines.(21) DhMab-1 can be used to analyze the expression and pathophysiological function of DGKη and represents a useful tool to study the molecular basis of various diseases.
Footnotes
Author Disclosure Statement
Y.K. received research funding from Ono Pharmaceutical Co., Ltd. The other authors have no conflict of interest.
Funding Information
This research was supported, in part, by AMED under Grant Nos. JP19am0401013 (Y.K.), JP19am0101078 (Y.K.), and JP19ae0101028 (Y.K.), and by JSPS KAKENHI Grant No. 17K07299 (M.K.K.) and Grant No. 19K07705 (Y.K.).