
Abstract
Anti-podoplanin (PDPN) monoclonal antibodies (mAbs) are needed as markers for lymphatic endothelial cells or type I alveolar cells in immunohistochemical analyses. We have developed anti-PDPN mAbs for many species, including humans, mice, rats, rabbits, dogs, cats, bovines, pigs, Tasmanian devils, alpacas, tigers, whales, goats, horses, and bears. This study develops and characterizes anti-sheep PDPN (sPDPN) mAbs using Cell-Based Immunization and Screening (CBIS) method. A RAP14 tag was added to the N-terminus of sPDPN, and an anti-RAP14 tag mAb (PMab-2) was used to measure the expression level of sPDPN in flow cytometry and Western blots. We immunized mice with sPDPN-overexpressing Chinese hamster ovary (CHO)-K1 (CHO/sPDPN) cells and screened mAbs against sPDPN using flow cytometry. Two of the mAbs, PMab-253 (immunoglobulin M [IgM], kappa) and PMab-260 (IgM, kappa), detected CHO/sPDPN cells specifically using flow cytometry and Western blots. Both PMab-253 and PMab-260 stained the renal glomerulus and Bowman's capsule, lymphatic endothelial cells of the lung and colon, and type I alveolar cells of the lung, suggesting PMab-253 and PMab-260, which were developed by CBIS method, can be applied to functional analyses of sPDPN. We also determined the binding epitope of PMab-253 and PMab-260 using flow cytometry. Analysis of sPDPN deletion mutants revealed that the N-terminus of the PMab-253 and PMab-260 epitope exists between amino acids 110 and 115 of sPDPN. Analysis of sPDPN point mutations revealed that the critical epitope of PMab-253 and PMab-260 includes Thr112 and Ser113 of sPDPN, indicating that the PMab-253 and PMab-260 epitope are independent of the platelet aggregation-stimulating (PLAG) domain or the PLAG-like domain of sPDPN.
Introduction
Sheep are used as experimental animals to study the immune system,(1–3) development of the blood–brain barrier,(4) and lymphopoiesis/lymphocyte recirculation,(5) among other subjects. However, detailed morphological studies of sheep lymphatic vessels have not been performed due to the lack of specific markers for lymphatic endothelial cells. Podoplanin (PDPN) is a type I transmembrane O-glycoprotein, which is expressed in renal podocytes, the lymphatic endothelial cells of every organ, and type I alveolar cells of the lung.(6,7) In morphological studies, PDPN is an important specific marker of lymphatic vessels.(7) PDPN possesses platelet aggregation-stimulating (PLAG) domains(8) and a PLAG-like domain,(9–13) which bind to C-type lectin-like receptor-2 (CLEC-2).(14) The PDPN/CLEC-2 interaction induces platelet aggregation and separation of embryonic blood/lymphatic vessels.(15)
We previously developed monoclonal antibodies (mAbs) against human,(16) mouse,(17) rat,(18) rabbit,(19) dog,(20) cat,(21) bovine,(22) pig,(23,24) Tasmanian devil,(25) alpaca,(26) tiger,(27) whale,(28) goat,(29,30) horse,(31,32) and bear(33,34) PDPNs using Cell-Based Immunization and Screening (CBIS) method.(35–37) These mAbs have all proven useful in flow cytometry, Western blots, and immunohistochemical analyses. In this study, we describe the development of anti-sheep PDPN (sPDPN) mAbs using CBIS to stain lymphatic endothelial cells by immunohistochemistry. We also determined the binding epitope of anti-sPDPN mAbs using flow cytometry.
Materials and Methods
Cell lines
Chinese hamster ovary (CHO)-K1 and P3X63Ag8U.1 (P3U1) cells were obtained from the American Type Culture Collection (Manassas, VA). Synthesized DNA (Eurofins Genomics KK, Tokyo, Japan) encoding sPDPN (Accession No.: XM_004013802.4) and an N-terminal RAP14 tag was subcloned into a pCAG-Neo vector (FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan). The RAP14 tag comprised 14 amino acids (aa), (DMVNPGLEDRIEDL) and is recognized by PMab-2 mAb(18) and LpMab-7.(38) sPDPN wild-type (WT) and mutation plasmids were transfected using Lipofectamine LTX with Plus Reagent (Thermo Fisher Scientific, Inc., Waltham, MA). Stable transfectants were selected by limiting dilution and cultivation in medium containing 0.5 mg/mL of G418 (Nacalai Tesque, Inc., Kyoto, Japan).
CHO-K1, P3U1, CHO/sPDPN,(39) CHO/human PDPN (hPDPN),(8) CHO/mouse PDPN (mPDPN),(8) CHO/rat PDPN (rPDPN),(18) CHO/rabbit PDPN (rabPDPN),(19) CHO/dog PDPN (dPDPN),(20) CHO/bovine PDPN (bovPDPN),(22) CHO/cat PDPN (cPDPN),(21) CHO/pig PDPN (pPDPN),(24) CHO/horse PDPN (horPDPN),(31) CHO/tiger PDPN (tigPDPN),(27) CHO/alpaca PDPN (aPDPN),(26) CHO/bear PDPN (bPDPN),(24) CHO/Tasmanian devil PDPN (tasPDPN),(24) CHO/goat PDPN (gPDPN),(27) and CHO/whale PDPN (wPDPN)(28) were cultured in a Roswell Park Memorial Institute (RPMI) 1640 medium (Nacalai Tesque, Inc.), supplemented with 10% heat-inactivated fetal bovine serum (Thermo Fisher Scientific, Inc.), 100 U/mL of penicillin, 100 μg/mL of streptomycin, and 0.25 μg/mL of amphotericin B (Nacalai Tesque, Inc.). Cells were grown in a humidified incubator at 37°C with an atmosphere of 5% CO2 and 95% air.
Animals
All animal experiments were performed according to relevant guidelines and regulations to minimize animal suffering and distress in the laboratory. Animal experiments were approved by the Animal Care and Use Committee of Tohoku University (Permit number: 2016MdA-153). Mice were monitored daily for health during the full 4-week duration of the experiment. A reduction of more than 25% of total body weight was defined as a humane endpoint. Mice were euthanized by cervical dislocation, and death was verified by respiratory and cardiac arrest. Female BALB/c mice (6 weeks old) were purchased from CLEA Japan (Tokyo, Japan). The animals were housed under specific pathogen-free conditions.
Production of sPDPN mutants
Deletion mutants of the sPDPN sequence were produced using a HotStar HiFidelity polymerase kit (Qiagen, Inc., Hilden, Germany) with oligonucleotides. Substitutions of amino acids with alanine in the sPDPN sequence were produced by QuikChange Lightning site-directed mutagenesis kits (Agilent Technologies, Inc., Santa Clara, CA). Polymerase chain reaction fragments bearing the desired mutations were inserted into the pCAG vector using the In-Fusion HD cloning kit (Takara Bio, Inc., Shiga, Japan).
Hybridoma production
To develop mAbs against sPDPN, we employed CBIS.(21,35–37) Briefly, three BALB/c mice were immunized with CHO/sPDPN cells (1 × 108) by the intraperitoneal route together with the Imject Alum (Thermo Fisher Scientific, Inc.). The procedure included three additional immunizations followed by a final booster intraperitoneal injection administered 2 days before the harvest of spleen cells. Harvested spleen cells were subsequently fused with P3U1 cells using PEG1500 (Roche Diagnostics, Indianapolis, IN), and the hybridomas were grown in an RPMI medium supplemented with hypoxanthine, aminopterin, and thymidine for selection (Thermo Fisher Scientific, Inc.). Culture supernatants were screened by flow cytometry.
Flow cytometry
Cells were harvested following a brief exposure to 0.25% trypsin and 1 mM ethylenediaminetetraacetic acid (Nacalai Tesque, Inc.). They were washed with 0.1% bovine serum albumin in phosphate-buffered saline (PBS) and treated with primary mAbs for 30 min at 4°C. The cells were then treated with Alexa Fluor 488-conjugated anti-mouse immunoglobulin G (IgG, 1:2000; Cell Signaling Technology, Inc., Danvers, MA) or Oregon green anti-rat IgG (1:2000; Thermo Fisher Scientific, Inc.). Fluorescence data were collected using the SA3800 Cell Analyzer (Sony Corp., Tokyo, Japan).
Determination of binding affinity by flow cytometry
CHO/sPDPN was suspended in 100 μL of serially diluted PMab-253 and PMab-260, and Alexa Fluor 488-conjugated anti-mouse IgG (1:200; Cell Signaling Technology, Inc.) was added. Fluorescence data were collected using the EC800 cell analyzer (Sony Corp.). The dissociation constant (KD) was calculated by fitting binding isotherms to built-in, one-site binding models in GraphPad PRISM 6 (GraphPad Software, Inc., La Jolla, CA).
Western blot analysis
Cell lysates (10 μg) were boiled in a sodium dodecyl sulfate sample buffer (Nacalai Tesque, Inc.). Proteins were then electrophoresed on 5%–20% polyacrylamide gels (FUJIFILM Wako Pure Chemical Corporation) and transferred onto polyvinylidene difluoride membranes (Merck KGaA, Darmstadt, Germany). After blocking with 4% skim milk (Nacalai Tesque, Inc.), the membranes were incubated with 5 μg/mL of PMab-253, 5 μg/mL of PMab-260, 1 μg/mL of anti-RAP14 tag (clone: PMab-2),(18) or 1 μg/mL of anti-β-actin (clone AC-15; Sigma-Aldrich Corp., St. Louis, MO) and incubated with peroxidase-conjugated anti-mouse immunoglobulins (Agilent Technologies, Inc.; diluted 1:2000). The membranes were finally developed using the Pierce Western Blotting Substrate Plus (Thermo Fisher Scientific, Inc.) or ImmunoStar LD (FUJIFILM Wako Pure Chemical Corporation) with a Sayaca-Imager (DRC Co. Ltd., Tokyo, Japan).
Immunohistochemical analyses
Normal sheep tissues from two sheep were collected after autopsies at Hokkaido University, fixed in 10% neutral-buffered formalin, and processed routinely to make formalin-fixed paraffin-embedded (FFPE) sections as in our previous study.(39) Histological sections 4-μm thick were directly autoclaved in a citrate buffer (pH 6.0; Nichirei Biosciences, Inc., Tokyo, Japan) for 20 min. After blocking with SuperBlock T20 (PBS) blocking buffer (Thermo Fisher Scientific, Inc.), the sections were incubated with PMab-253 and PMab-260 (5 μg/mL) for 1 h at room temperature and treated using an Envision+ kit (Agilent Technologies, Inc.) for 30 min. Color was developed using 3,3′-diaminobenzidine tetrahydrochloride (Agilent Technologies, Inc.) for 2 min, and counterstaining was performed with hematoxylin (FUJIFILM Wako Pure Chemical Corporation).
Results
Establishment of anti-sPDPN mAbs
To produce anti-sPDPN mAbs, we employed CBIS, using stable transfectants for both immunization and flow cytometric screening. Three mice were immunized with CHO/sPDPN cells, which overexpress sPDPN. Hybridomas were seeded into 96-well plates, and CHO/sPDPN-positive and CHO-K1-negative wells were selected. PMab-253 (immunoglobulin M [IgM], kappa) and PMab-260 (IgM, kappa) were selected using Western blotting and immunohistochemistry.
Flow cytometric analyses
We performed flow cytometry using PMab-253 and PMab-260 against CHO/sPDPN and CHO-K1. Both PMab-253 and PMab-260 recognized CHO/sPDPN, but not CHO-K1 (Fig. 1A, B). As a positive control, an anti-RAP14 tag mAb (PMab-2) detected CHO/sPDPN. The results indicate that PMab-253 and PMab-260 can detect sPDPN in flow cytometry analysis.
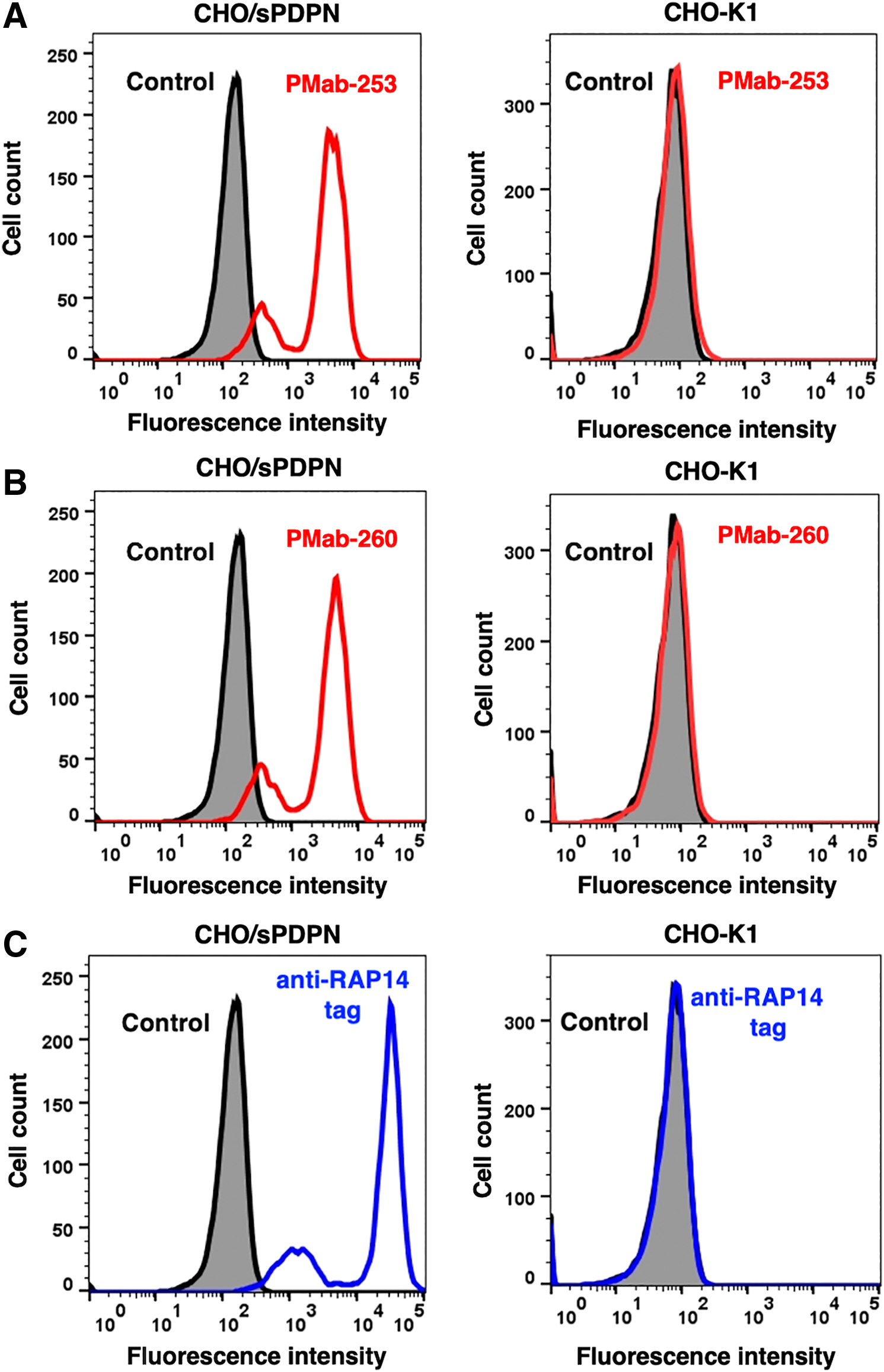
Detection of sPDPN by flow cytometry using anti-sPDPN mAbs. CHO/sPDPN and CHO-K1 cells were treated with PMab-253
Next, the cross-reaction of PMab-253 and PMab-260 with the other PDPNs, which are overexpressed in CHO-K1 cells, was investigated. PMab-253 cross-reacted with hPDPN, bovPDPN, cPDPN, gPDPN, tigPDPN, and wPDPN (Fig. 2A). In contrast, PMab-260 cross-reacted with hPDPN, bovPDPN, gPDPN, and wPDPN (Fig. 2B).

Cross-reaction of anti-sPDPN mAbs with PDPNs of other species by flow cytometry. CHO-K1 cells transfected with PDPNs of other species were treated with PMab-253
We then assessed the apparent binding affinity of PMab-253 and PMab-260 with CHO/sPDPN using flow cytometry. The KD of PMab-253 and PMab-260 for CHO/sPDPN was 2.6 × 10−8 M and 2.7 × 10−8 M, respectively (Fig. 3), indicating that PMab-253 and PMab-260 possess moderate affinity for CHO/sPDPN cells.
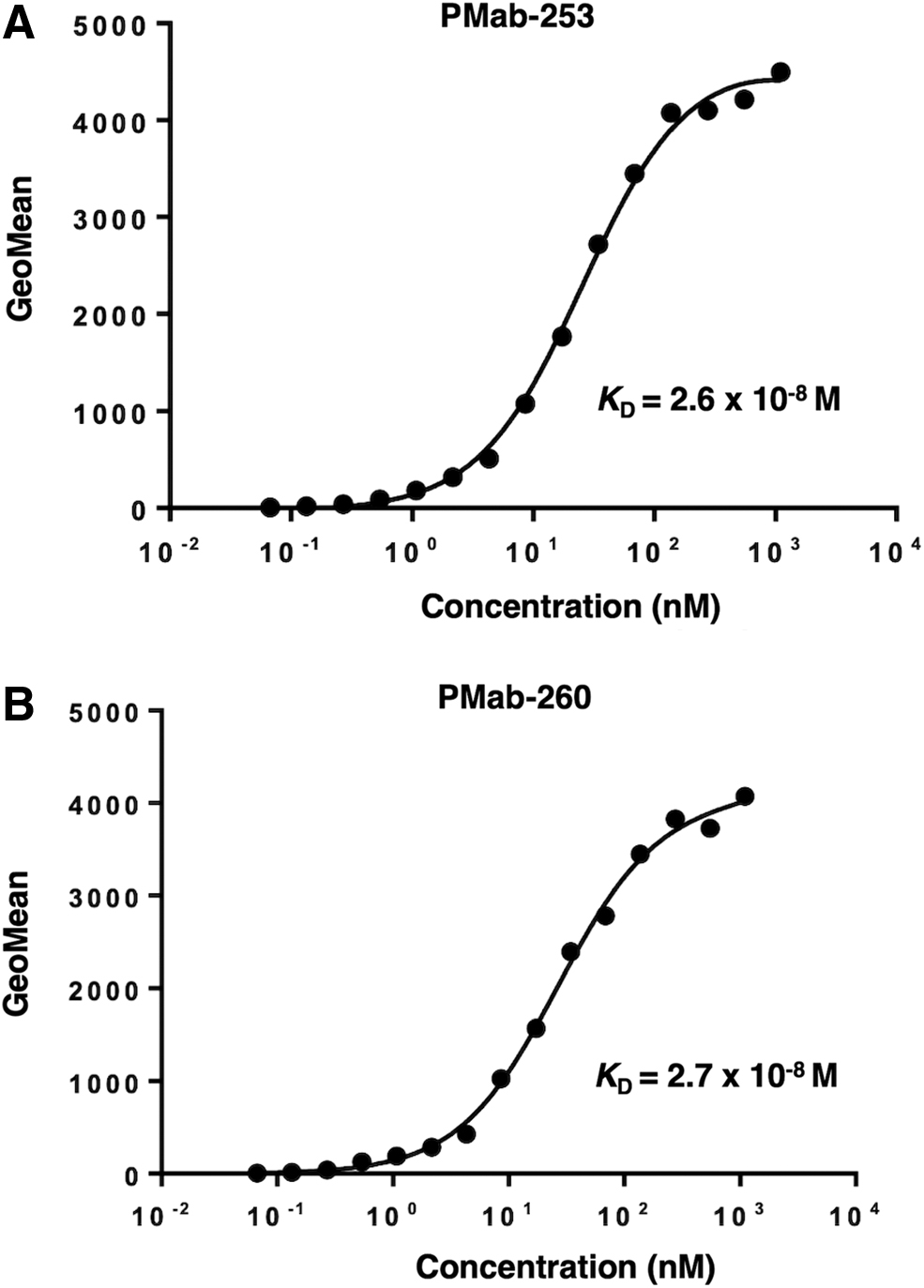
Determination of binding affinity of anti-sPDPN mAbs. CHO/sPDPN was suspended in 100 μL of serially diluted PMab-253
Western blot analyses
We next investigated whether PMab-253 and PMab-260 can be applied to Western blot analysis by using cell lysates of CHO-K1 or CHO/sPDPN. As shown in Figure 4, PMab-253 and PMab-260 detected sPDPN as a 55-kDa band (N-terminal RAP14 tag + sPDPN) in CHO/sPDPN cell lysates, but these antibodies did not detect any band from CHO-K1 cells. Anti-RAP14 tag (PMab-2) as a positive control also detected a band of the same molecular weight from CHO/sPDPN. These results indicate that PMab-253 and PMab-260 can detect sPDPN in Western blot analysis.
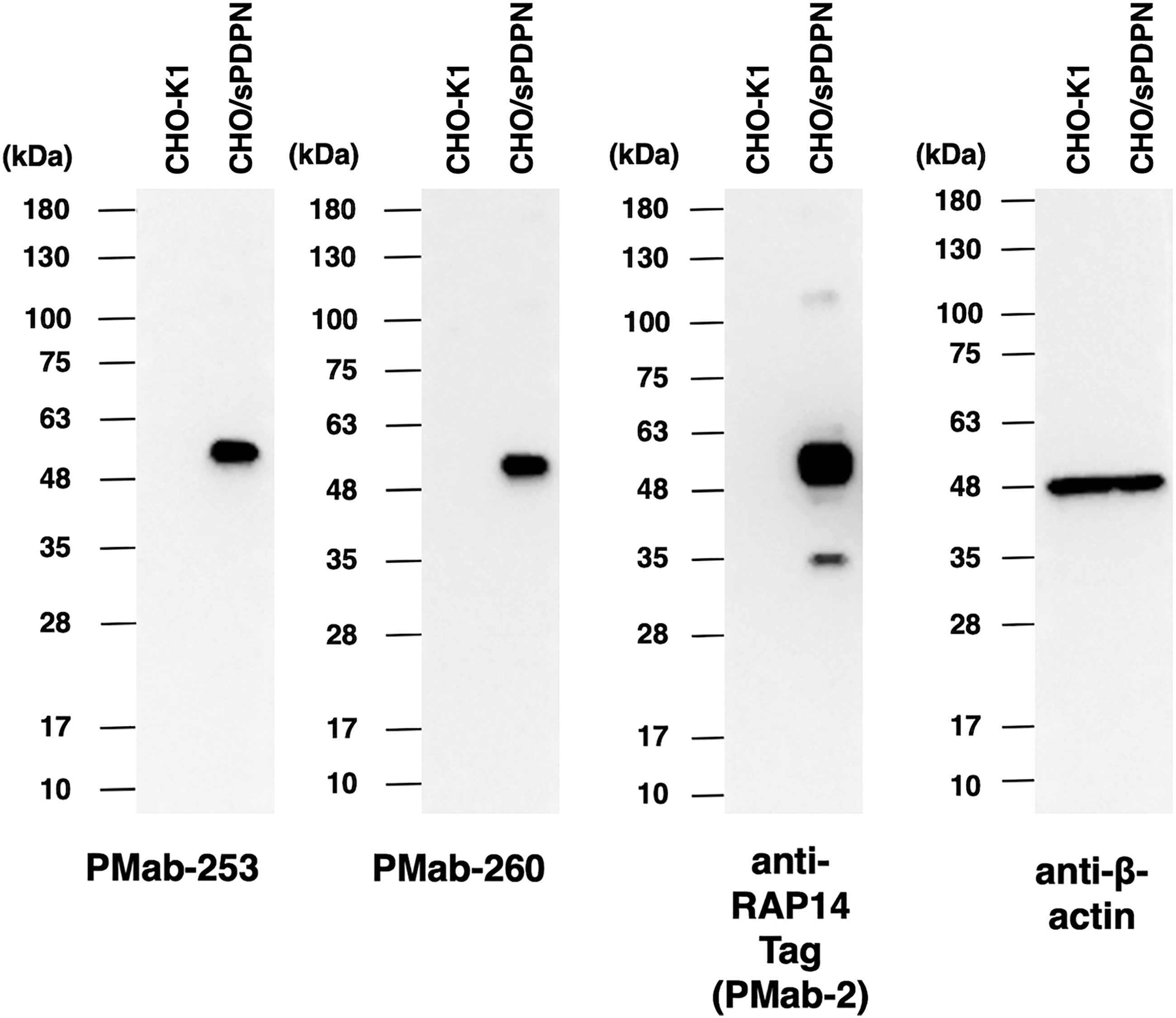
Western blot analysis. Cell lysates of CHO-K1 and CHO/sPDPN (10 μg) were electrophoresed and transferred onto polyvinylidene fluoride membranes. The membranes were incubated with 5 μg/mL of PMab-253, 5 μg/mL of PMab-260, 1 μg/mL of anti-RAP14 tag (PMab-2), and 1 μg/mL of anti-β-actin and subsequently with peroxidase-conjugated anti-mouse immunoglobulins.
Immunohistochemical analyses
We next investigated whether PMab-253 and PMab-260 can be applied to immunohistochemical analysis using FFPE sheep sections. For this assessment, we selected normal sheep lung, kidney, and colon tissues, all of which reportedly express PDPN in other species, including humans,(16) mice,(17) rats,(18) rabbits,(19) dogs,(20) cats,(21) bovines,(22) pigs,(23, 24) Tasmanian devils,(25) alpacas,(26) tigers,(27) whales,(28) goats,(29,30) horses,(31,32) and bears.(33,34) As shown in Figure 5A and B, PMab-253 stained the renal glomerulus and Bowman's capsule. Lymphatic endothelial cells (red arrow) of the colon were readily detected by PMab-253 (Fig. 5C, D). Furthermore, PMab-253 strongly stained type I alveolar cells and lymphatic endothelial cells (red arrow) of the lung (Fig. 5E, F). Similarly, PMab-260 strongly stained the renal glomerulus (Fig. 6A, B), lymphatic endothelial cells (red arrow) of colon (Fig. 6C, D), and type I alveolar cells and lymphatic endothelial cells (red arrow) of the lung (Fig. 6E, F).

Immunohistochemical analyses of sheep tissues using PMab-253. Formalin-fixed paraffin-embedded tissue sections of sheep kidneys
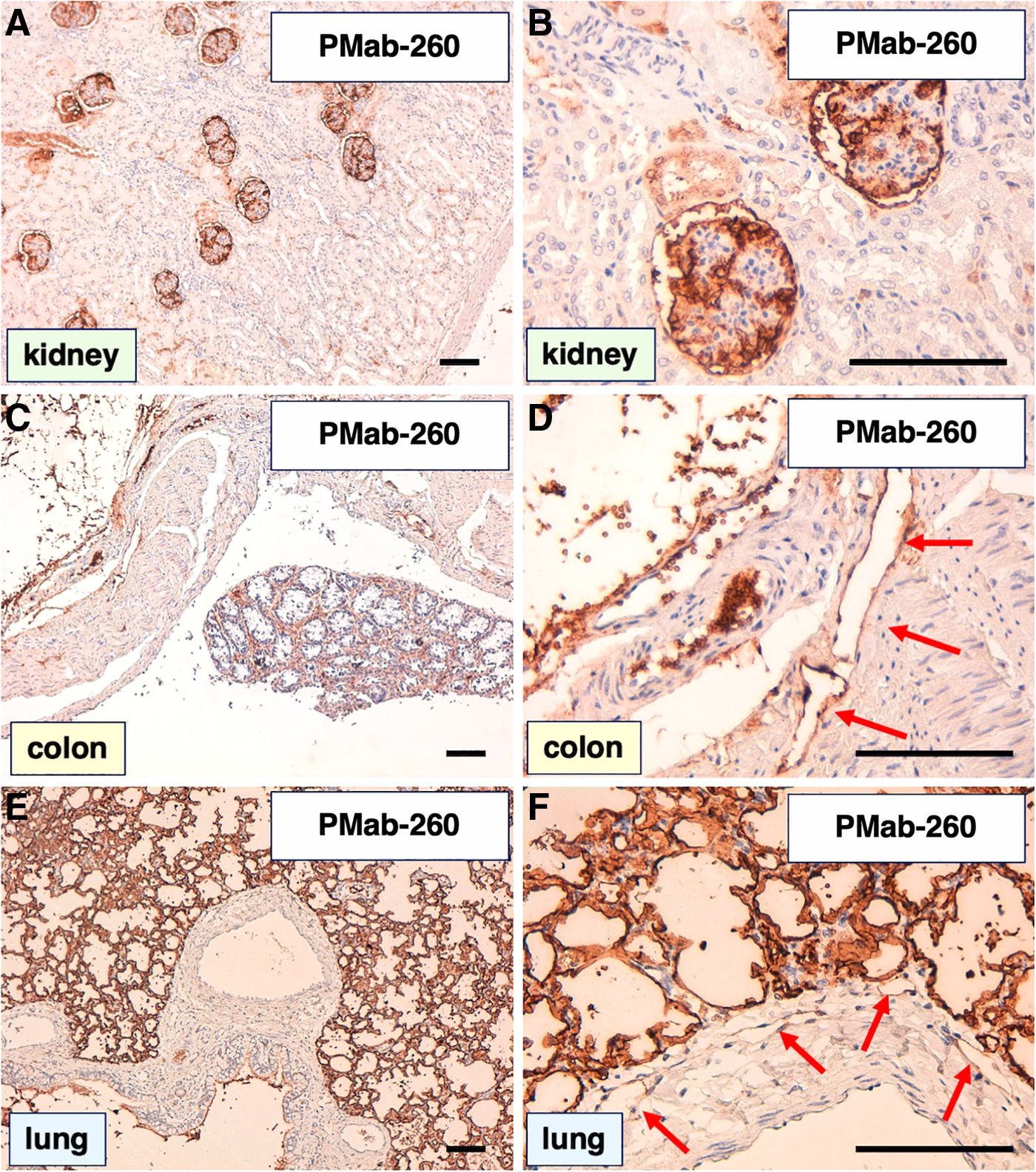
Immunohistochemical analyses of sheep tissues using PMab-260. Formalin-fixed paraffin-embedded tissue sections of sheep kidneys
Epitope mapping
Eleven deletion mutants of sPDPN in CHO-K1 cells were produced as follows: dN30, aa 30–168; dN40, aa 40–168; dN50, aa 50–168; dN60, aa 60–168; dN70, aa 70–168; dN80, aa 80–168; dN100, aa 100–168; dN105, aa 105–168; dN110, aa 110–168; dN115, aa 115–168; dN120, aa 120–168; or WT sPDPN, aa 27–168. All deletion mutants and WTs containing the N-terminal RAP14 tag were recognized by PMab-2 (an anti-RAP14 tag mAb), indicating that the expression level of each construct was high (Fig. 7A). PMab-253 recognized dN30, dN40, dN50, dN60, dN70, dN80, dN100, dN105, and dN110, but did not react with dN115 and dN120 (Fig. 7B), suggesting that the N-terminus of the PMab-253 epitope is between aa 110 and 115 of sPDPN. Similarly, PMab-260 recognized dN30, dN40, dN50, dN60, dN70, dN80, dN100, dN105, and dN110, but did not react with dN115 and dN120 (Fig. 7C), suggesting that the N-terminus of the PMab-260 epitope is also between aa 110 and 115 of sPDPN.
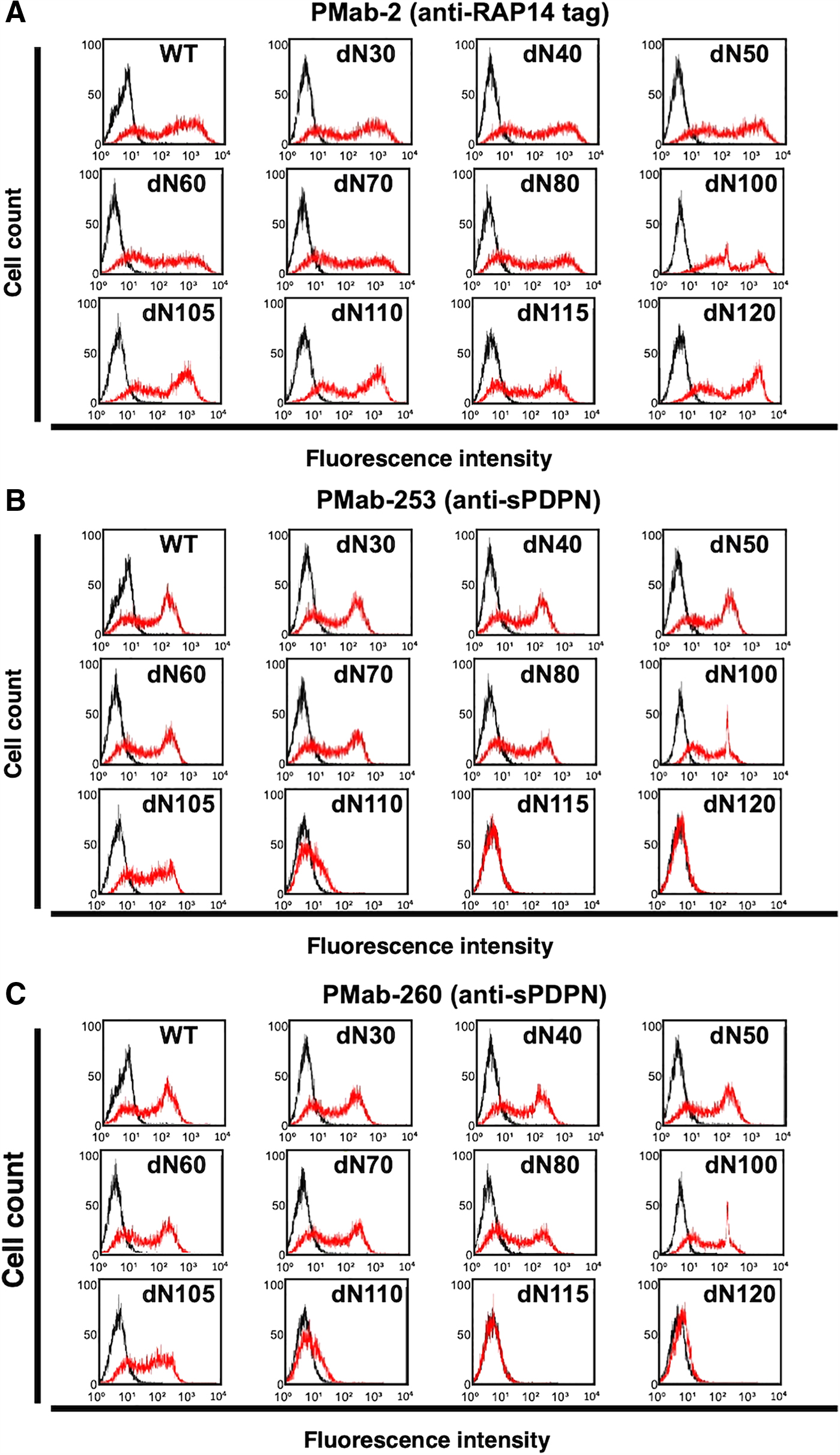
Epitope mapping of PMab-253 and PMab-260 using deletion mutations.
Next, we obtained a series of point mutants of sPDPN: V105A, H106A, T107A, H108A, V109A, P110A, A111G, T112A, S113A, H114A, S115A, and Q116A. PMab-2 reacted with all point mutants (Fig. 8A), but PMab-253 reacted less with T112A and S113A (Fig. 8B), indicating that Thr112 and Ser113 of sPDPN are essential for PMab-253 binding. PMab-260 reacted with T112A weakly, and did not react with S113A (Fig. 8C), also indicating that Thr112 and Ser113 of sPDPN are essential for PMab-260 binding. The epitope of PMab-253 and PMab-260 is illustrated in Figure 9A and B, respectively.

Epitope mapping of PMab-253 and PMab-260 using point mutations.

Schematic illustration of the epitope recognized by PMab-253 and PMab-260.
Discussion
We previously investigated possible cross-reactions between anti-PDPN mAbs and sPDPN.(39) Flow cytometry analysis demonstrated that an anti-bovPDPN mAb (clone PMab-44)(22) reacted with CHO/sPDPN. Other anti-PDPN mAbs such as anti-mPDPN mAb (PMab-1),(17) anti-rPDPN mAb (PMab-2),(18) anti-rabPDPN mAb (PMab-32),(19) anti-dPDPN mAb (PMab-38 and PMab-48), and anti-cPDPN mAb (PMab-52)(21) did not react with CHO/sPDPN, indicating that only PMab-44 was useful for the detection of sPDPN. Expression of sPDPN was then investigated by immunohistochemical analyses using PMab-44. Although type I alveolar cells of sheep lungs were stained by PMab-44, the renal glomerulus and lymphatic endothelial cells were not stained by PMab-44.(39) As a result, novel anti-sPDPN mAbs for detecting lymphatic endothelial cells by immunohistochemical analysis are needed.
In this study, we immunized mice with CHO/sPDPN cells, performed the first screening using only flow cytometry, and established two mAbs (PMab-253 and PMab-260). Although our previous study showed that PMab-44 stained only type I alveolar cells and not lymphatic endothelial cells from sheep organs, PMab-253 and PMab-260 strongly stained lymphatic endothelial cells of the lung or colon (Figs. 5 and 6). Our recent study showed that one of the anti-bPDPN mAbs (clone PMab-241) reacted with lymphatic endothelial cells, but not with type I alveolar cells of bear lungs.(34) The other anti-bPDPN mAb (clone PMab-247) reacted not only with lymphatic endothelial cells but also type I alveolar cells and renal podocytes,(33) suggesting that the binding epitope of each mAb is critical to distinguishing PDPN-expressing lymphatic endothelial cells from other PDPN-expressing tissues. We therefore determined the binding epitope of PMab-253 and PMab-260. Using a series of point mutants of sPDPN, Thr112 and Ser113 of sPDPN were determined to be essential for PMab-253 and PMab-260 binding. Until now, we have not found the binding epitope of anti-PDPN mAbs between aa 100 and aa 120 near the transmembrane domain of PDPN, indicating that the epitope of PMab-253/PMab-260 is novel. PMab-253 and PMab-260 may be advantageous for detailed morphological studies of sheep lymphatic vessels.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported, in part, by Japan Agency for Medical Research and Development (AMED) under Grant Numbers: JP20am0401013 (Y.K.), JP20am0101078 (Y.K.), and JP20ae0101028 (Y.K.), and by Japan Society for the Promotion of Science (JSPS) KAKENHI Grant Numbers: 17K07299 (M.K.K.), 19K07705 (Y.K.), and 20K16322 (M.S.).