
Abstract
During preclinical studies, there is a great need to develop monoclonal antibodies (mAbs) that are specific to human immunoglobulin (IgG), without binding to monkey IgG, to detect therapeutic human mAb in non-human primates. We took advantage of the latest rabbit B cell cloning technology to develop six unique rabbit anti-human IgG mAb clones for this purpose. These clones are capable of binding to both human IgG and Fab with high affinity without nonspecific binding to cynomolgus monkey IgG. These clones have been evaluated as a generic capture reagent for the detection of human IgG and Fab, in the presence of cynomolgus monkey serum, by Gyrolab™ immunoassay. They may be used in singlet or as pairs for the detection of human IgG, in any host animal, to meet the need for therapeutic mAb development in preclinical studies.
Introduction
In the field of modern medicine, therapeutic monoclonal antibodies (mAbs) have become one of the fastest growing classes of drugs in recent years and are approved for the treatment of a wide range of indications, from cancer to autoimmune diseases.(1) Factors related to antigen expression, target pharmacology, and antibody-effector functions can contribute to the adverse event profiles observed with mAbs. Thus, the effective translation of information gained from preclinical research and safety studies, into clinical development, is a crucial step for successful development of mAbs.(2)
Preclinical pharmacokinetic characterization of these therapeutic mAbs often has to be performed in non-human primates to prove efficacy and safety before the initiation of clinical studies. The cynomolgus monkey is a preferred non-human primate for preclinical studies with therapeutic antibodies because it often provides a sufficient level of cross-reactivity with the target of the therapeutic antibody.(3) However, the immunoglobulins (IgGs) of the cynomolgus monkey also show a high sequence homology with the IgGs of humans—and due to the lack of high-quality reagent antibodies that can distinguish the human therapeutic molecule from cynomolgus monkey IgG in the serum—this high level of protein sequence homology for IgG constitutes a significant challenge for the bioanalytical measurement of human therapeutic antibodies in non-human primate serum samples.(4)
Traditionally, the majority of mAbs have been generated in mice; however, the use of rabbit mAbs has been increasing in recent years.(5) In general, rabbit mAbs offer several advantages over traditional mouse mAbs, such as higher affinity and specificity than mouse antibodies, as well as more diverse epitope recognition than mouse antibodies.(5,6) The rabbit's immune system is evolutionarily distinct from that of rodents and it uses different mechanisms to generate, diversify, and optimize the affinity of the antibodies it produces. In addition, the rabbit's immune system can recognize smaller-sized epitopes that are not immunogenic in mice, while maintaining the ability to produce strong immune responses.(6,7) Therefore, generating mAbs with rabbits instead of mice, particularly for reagent purposes, has become increasingly popular.(8)
Although hybridoma screening and display methodologies have been used in rabbit mAb development, they both have some drawbacks: the hybridoma technology has a low efficiency of cell fusion, while display methods result in the loss of natural cognate pairing between heavy and light chains.(6) To overcome these issues, a single B cell-based antibody gene cloning technology (or single B cell cloning) has recently been developed.(8,9) Briefly, the single B cell cloning consists of the following steps: (1) isolation of specific single B cells from peripheral blood or from lymphoid tissues by antigen-based fluorescent-activated cell sorting, (2) growth and expansion of the single B cell for 2 weeks, (3) amplification and sequencing of the antibody genes by reverse transcription-polymerase chain reaction (RT-PCR) with antibody-specific primers, (4) cloning of antibody genes into an expression vector and producing recombinant mAb in mammalian cell systems (e.g., HEK 293 and CHO cells), and (5) purification of the resulting recombinant mAb and evaluation by enzyme-linked immunosorbent assay (ELISA) and other in vitro assays.(6) Over the years, efforts have been made to optimize each step of this process to improve the efficiency and success rate.
In this work, a human IgG Fab was used to immunize rabbits. The splenocytes were isolated, and flow cytometry was used to sort human IgG-specific B cells. Using cynomolgus monkey IgG as a counter screening agent, we obtained six human IgG- and Fab-specific mAb clones. These clones demonstrated better binding affinity and targeted different epitopes from the only commercially available anti-hIgG mAb of mouse-origin clone MCA5748G. Each of the rabbit anti-hIgG mAb clones obtained from the single B cell cloning process has been evaluated by Gyrolab™ assays and found to be suitable for use as capture reagent in generic pharmacokinetic assays in the presence of cynomolgus monkey serum.
Materials and Methods
Chemicals and reagents
The antibodies used in this work were a humanized therapeutic mAb IgG1 (hIgG1), a humanized development candidate mAb IgG4 (hIgG4), and an internal human Fab (hFab) research reagent. Cynomolgus monkey IgG (cynoIgG) was purified from cynomolgus monkey serum, which was purchased from Innovative Research (Novi, MI), by protein A affinity purification. The mouse anti-human IgG mAb MCA5748G(4) was purchased from BioRad Laboratories (Hercules, CA). Cell culture medium and phosphate-buffered saline (PBS) were purchased from ThermoFisher Scientific (Waltham, MA). All other chemicals were of analytical grade.
ELISA for determination of binding specificity
An ELISA was used to evaluate the specificity of the antibody clones for binding to human IgG using cynoIgG as a control. The ELISA was performed at room temperature on microtiter plates from ThermoFisher Scientific, which were first coated with hIgG1, hIgG4, or cynoIgG in PBS for 1 hour. After washing thrice with PBS-polysorbate 20 (Tween 20), the plate was blocked with PBS/3% bovine serum albumin for 1 hour. The plate was then washed again and incubated with anti-human IgG antibody clones for 1 hour. After another washing step, the bound antibodies were detected by horseradish peroxidase -conjugated anti-rabbit IgG antibody from Southern Biotech (Birmingham, AL), according to the manufacturer's instructions.
Biacore assay for determination of binding specificity and kinetics
The specificity of each rabbit anti-human IgG mAb was evaluated in a second assay system.(10) These experiments were performed with the Biacore® T200 instrument (Biacore, Uppsala, Sweden) using Streptavidin or CM5 sensor chips from Biacore. Coating of an antibody to the streptavidin chip was achieved by injecting biotinylated target antibody, which was amine coupled using the EZ-Link™ Amine-PEG11-Biotin reagent from ThermoFisher Scientific following the manufacturer's manual. For the CM5 chip, the target antigen or antibody was coupled to the chip surface, by standard amine coupling, using the amine coupling kit from Biacore. Unless otherwise stated, all binding and kinetics assays were performed in HBS-EP+ buffer (0.01 M HEPES pH 7.4, 0.15 M NaCl, 3 mM EDTA, and 0.005% v/v Surfactant P20) at 25°C. The dissociation constant values (KD) were calculated with a 1:1 Langmuir fitting model using BIAevaluation software V4.1 from Biacore.
Rabbit immunization and B cell cloning
An anti-human CD38 Fab, as described by Wu et al.,(11) was used to immunize two rabbits with a total of five antigen injections. The primary injection used complete Freund's adjuvant (CFA) and the four boosts used incomplete Freund's adjuvant (IFA). The CFA and IFA were both from ThermoFisher Scientific. Serum titers were monitored by ELISA using the antigen protein. The rabbit with the higher ELISA titer was chosen for splenectomy. The U.S. federal Animal Welfare Act and the Public Health Service Policy on Humane Care and Use of Laboratory Animals (PHS Policy) guidelines for the care and use of laboratory animals were followed.
For B cell isolation, fresh splenocytes were isolated from the spleen. Approximately 1.2 × 108 splenocytes were cultured overnight in a special B cell medium customized by Yurogen (Worcester, MA) before sorting. Splenocytes were processed using the SMab™ platform at Yurogen to enrich antigen-recognizing B cells. The antigen-sorted B cells were seeded and cultured in a 96-well plate, with one cell per well, for 10–14 days.
Antigen-recognizing B cell clones were identified and confirmed using hIgG4-coated direct ELISA, and purified cynoIgG was used in ELISA for counter screening. Antigen-specific B cell clones were ranked and selected according to their positive/negative ELISA signal ratio and their heavy- and light-chain IgG coding sequences were amplified by RT-PCR. The heavy- and light-chain PCR products were combined and used to transfect HEK293F cells directly. Transiently expressed recombinant rabbit IgG clones were then further confirmed for specific binding to hIgG1, hIgG4, and hFab by ELISA and Biacore binding assays. Upon confirming specific binding to hIgG1, hIgG4, and hFab, the PCR products from the selected positive B cells were cloned into a mammalian expression vector for scale-up antibody production in HEK293F cells. Recombinant rabbit mAb clones produced by HEK293F transfection were purified using protein A chromatography for further evaluation.
Gyrolab assay
Gyrolab xPlore, BioAffy 1000 nL CDs, and Rexxip A and Rexxip F Buffer from Gyros Protein Technologies (Uppsala Sweden) were used for all experiments.(12) Biotinylated capture antibodies were diluted to 0.1–0.2 μg/μL in Rexxip A Buffer and flowed over the streptavidin bead column within the microstructure of the BioAffy CDs. The standard curve and quality control (QC) samples were prepared by spiking the hIgG4 or hFab in Rexxip A Buffer containing various amounts of cynomolgus monkey serum. The standard curve, QC samples, mock samples, and assay reagents were added to 96-well plates and loaded onto the Gyrolab instrument. A single replicate of the standard curve, QC samples, and mock samples was added into two BioAffy CD microstructures by the Gyros instrument and then flowed over the bead column. Alexa Fluor 647-labeled goat anti-human IgG (Fc) antibody purchased from Southern Biotech (Birmingham, AL) was used as the detection reagent at a concentration of 2μg/mL in Rexxip F buffer. A wash solution of PBS with 0.01% v/v Tween-20 was flowed over the columns before each run to prewet the streptavidin beads, and after each step in the assay, to rinse away any unbound reagents. Sample concentrations were determined with the data acquisition at a 1% level of photomultiplication. The Gyrolab Evaluator software was used to analyze results with a 5-parameter fit and 1/Y2 weighting as directed by the manufacture.
Results and Discussion
Isolation of human IgG-specific clones
As described in Materials and Methods section, a human anti-CD38 Fab was used to immunize rabbits and the resultant splenocytes were sorted to isolate cells able to bind biotinylated hIgG1. A total of 530 primary B cells, which were able to bind to hIgG1, were single-cell seeded into 96-well plates and grown for 2 weeks. The B cell culture medium, which contained monoclonal rabbit IgG antibody, was screened by direct ELISA using hIgG4, hIgG1, and cynoIgG (data not shown). Based on the ELISA hIgG1/cynoIgG and hIgG4/cynoIgG signal values, 17 clones, which had both hIgG1 and hIgG4 OD450 >0.9, and CynoIgG OD450 <0.2, were selected (Supplementary Table S1), and their antibody coding sequences were amplified by RT-PCR. For 11 of the 17 clones, we were able to obtain PCR products for both heavy and light chains. The heavy- and light-chain PCR products from each clone were then combined at a 1:1 molar ratio and used to transfect HEK293F cells directly. Cell culture medium from the transfected HEK293F cells was confirmed by the same ELISA screen assay. As shown in Figure 1A,6 of the 11 clones exhibited good hIgG4/cynoIgG and hIgG1/cynoIgG ELISA signal ratio ranging from 4.94 to 14.99. These six clones were further analyzed for direct binding to hIgG1, hIgG4, hFab, and cynoIgG by Biacore as shown in Figure 1B. All six clones showed various levels of biding to hIgG1, hIgG4, and hFab, but not to cynoIgG, indicating that these clones were specific to human IgGs and Fab, but not to cynoIgG.
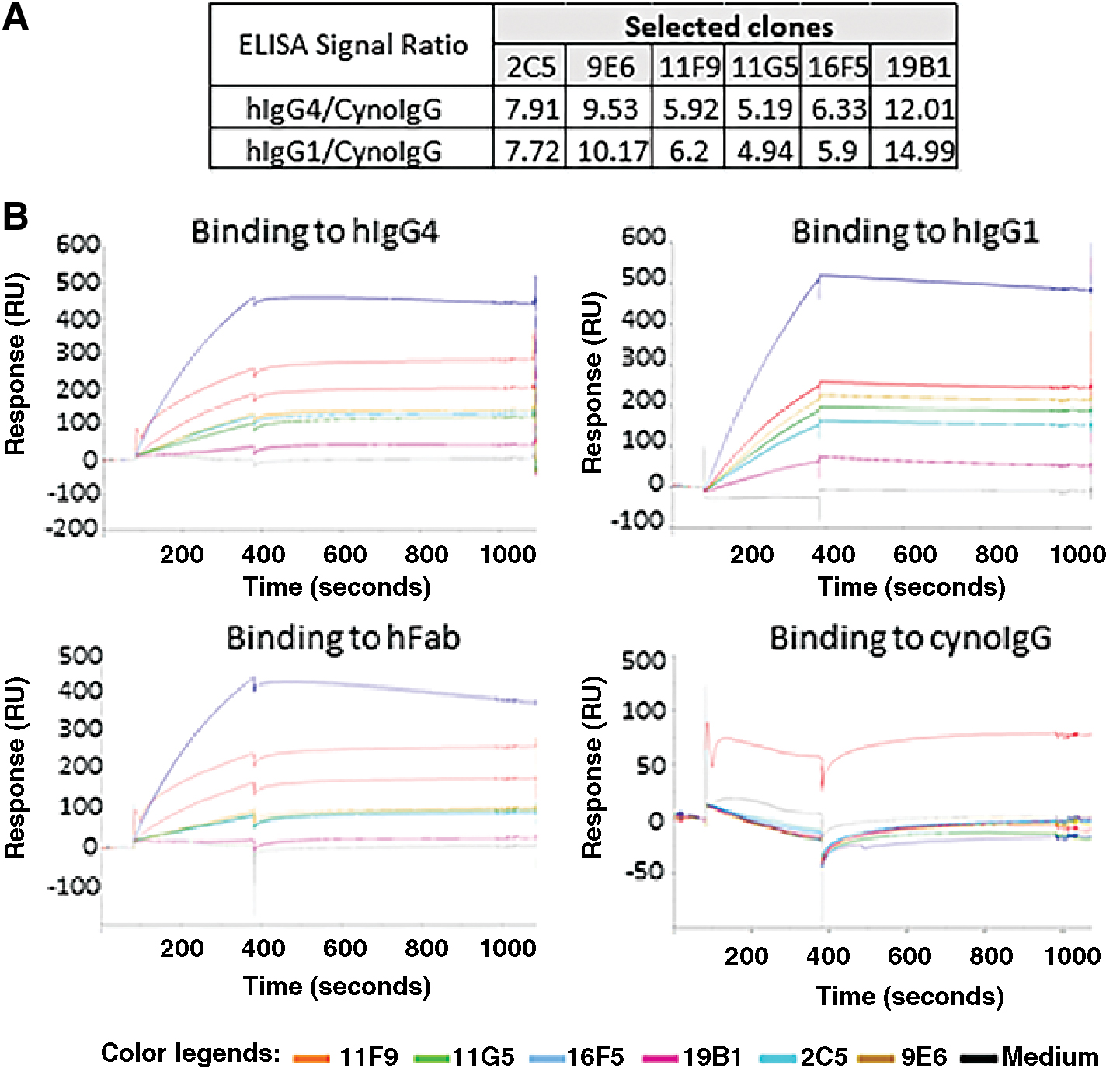
Selection of six rabbit recombinant antibody clones with specific binding to whole human IgG and Fab, but not to cynomolgus monkey IgG. Heavy- and light-chain coding sequences of selected B cell clones were amplified and transfected into HEK 294 cells. Cultural medium was harvested after 4 days of transfection and used in ELISA and Biacore assays.
Sequences of rabbit anti-hIgG clones
To determine the uniqueness of the six clones, heavy- and light-chain PCR products were subjected to DNA sequencing. The deduced amino acid sequences of the variable regions for the heavy and light chains, respectively, were aligned using the EMBL-EBI web-based Clustal Omega.(13) As shown in Figure 2, all six clones possess unique AA sequences for both heavy and light chains. The phylogenetic trees, based on Clustal Omega, were constructed to visualize relationship among these clones. Notably, clone 16F5 is highly divergent from other five clones in both heavy and light chains, whereas clone 11F9 is highly divergent in the heavy chain, but not in the light chain. Clones 9E6, 11G5, 2C5, and 19B1, on the other hand, were closely related to each other as demonstrated by their heavy-chain variable sequences.

Heavy- and light-chain variable region multiple alignments using Clustal Omega.
Binding affinity and epitope of the rabbit anti-hIgG mAb clones
The amplified PCR products containing the heavy- and light-chain antibody coding sequences were cloned into expression vectors and used for transient transfection and antibody purification. Using amine coupling, a mouse anti-rabbit mAb was directly immobilized on CM5 chip.(10) The purified rabbit mAb clones were then injected, followed by flow over 1:2 serially diluted hFab, hIgG1, hIgG4, and cynoIgG, ranging from 80 to 0 nM. As illustrated in Table 1, all the clones exhibited greater than subnanomolar binding affinity (KD) to hIgG1, hIgG4, and hFab, but not to cynoIgG. Under the same conditions, the commercial control clone MCA5748 was able to bind to hIgG1 and hIgG4, but not hFab and cynoIgG.
Biacore Kinetics of Selected Six Rabbit Anti-hIgG Clones Binding to Different Types Antibodies
To determine if the six rabbit anti-hIgG clones bound to different epitopes from the commercially available clone MCA5748G, we biotinylated MCA5748G, 19B1, 11F9, and a mouse anti-hIgG mAb as the assay control. These biotinylated antibodies were then injected into the different flow cells of a Biacore streptavidin chip to reach a density range between 500 and 600 response units, followed by flow over the 1:2 serially diluted hIgG1 (Isa, Isatuximab marketed by Sanofi) antibody, ranging from 80 to 0 nM, in the presence of 240 nm of different rabbit mAb clones as the competitor. As shown in Figure 3, when MCA5748G was used as the capture antibody on the chip surface, competition was only seen with mouse clone MAC5748G itself, indicating that this commercial mouse anti-hIgG clone MAC5748G does not compete with other clones; thus, it has a different binding epitope from all six of the rabbit anti-hIgG clones we developed (for additional data, please see Supplementary Fig. S1).
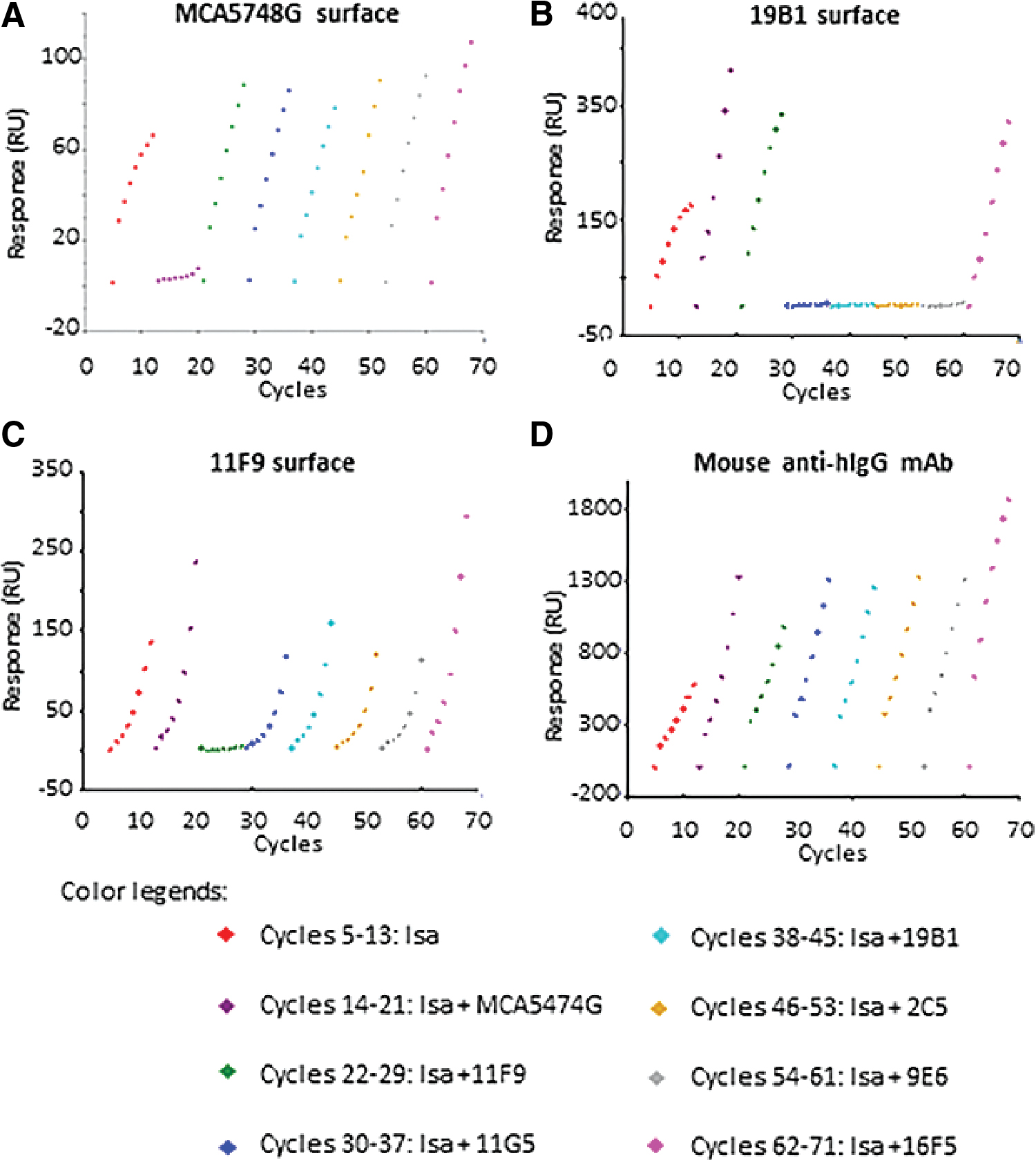
The binding level of Biacore competition assay. MCA5748G, 19B1, 11F9, and mouse anti-hIgG mAb
When the rabbit anti-hIgG clone 19B1 was used as the capture antibody on the Biacore chip's surface, competition was seen with clones 11G5, 19B1, 2C5, and 9E6. Furthermore, when clone 11F9 was captured on the chip's surface, competition was only seen with clone 11F9 itself. Clone 16F5 did not compete with any of the other clones. These results indicate that clones 16F5 and 11F9 both have a unique epitope, whereas the clones 11G5, 19B1, 2C5, and 9E6 share the same epitope. Paratopes in the complementarity-determining regions determine antibody binding epitopes and the heavy chain plays a dominant role in antigen binding,(14) and epitope classification of the six rabbit anti-hIgG clones correlates well with the phylogenetic tree's relationship.
Although it is not known which amino acid residues of the hIgGs make up each of the epitopes, or whether these epitopes are linear or conformational, because a Fab was used as the immunogen to generate the rabbit anti-hIgG mAb clones, all of them should bind within the Fab region of the human IgG molecule. Since different human IgG subtypes (IgG1, IgG2, IgG3, and IgG4) are classified by their Fc sequences—which are not part of the Fab region—each of the rabbit anti-hIgG mAb clones should have the ability to recognize all subtypes of human IgG. This is consistent with data from additional Biacore experiments, which indicate that all six of the rabbit anti-hIgG clones are capable of binding to all hIgGs and Fabs tested to date (10 different human IgGs and 2 Fabs, data not shown). The ability to use these rabbit anti-hIgG mAbs in preclinical studies and/or in process development, for the detection of the different subtypes of therapeutic human IgG (IgG1, IgG2, and IgG4), is a significant advantage.
Specific detection of human IgG by rabbit anti-hIgG mAb clones by Gyrolab assay
To determine if the rabbit anti-hIgG mAbs can detect human IgG molecules in the presence of monkey serum, biotinylated clone 16F6 was first used as the capture reagent. The assay matrixes used were Gyrolab Rexxip A Buffer mixed with 0%, 4%, 10%, and 25% cynomolgus monkey serum, respectively. A human IgG4 antibody was used to prepare the standard curve in assay matrix. As shown in Figure 4A, with a curve range of 1200–0.3 ng/mL, no matrix effect was observed with cynomolgus monkey serum concentrations between 0% and 4%. When the assay matrix contained 10% and 25% of the cynomolgus monkey serum, some minor background signal was observed on the lower half of the standard curve. To further evaluate recovery of hIgG, hIgG4 was spiked into the assay matrix containing 4% cynomolgus monkey serum to generate QC samples at 750, 40, and 0.6 ng/mL of hIgG4 and assessed in the assay. As shown in Figures 4B, in the 4% cynomolgus serum matrix, back-fit values of the standard curve and interpolated values of the QCs showed minimal bias (<20% bias, data not shown).

Gyrolab™ immunoassay using clone 16F5 as the capture reagent. Clone 16F5 was biotinylated and used as the capture reagent. The human IgG4 antibody (hIgG4) served as the assay standard, was diluted 1 to 4 at the range of 1200–0.30 ng/mL. Alexa Fluor 647-conjugated goat anti-human IgG was used for detection.
We further tested the suitability of the other rabbit anti-hIgG clones to be used as the capture reagent in Gyrolab assay. All six rabbit anti-hIgG clones, along with the commercial mouse anti-hIgG mAb MCA5748G clone, were biotinylated and used at a similar concentration in the Gyrolab assay. The Gyrolab generic PK assay capture reagent was used as control. As shown in Figure 5A, when the whole antibody molecule hIgG4 was used as the standard (same range from 1200 to 0.3 ng/mL), all of the capture reagents generated concentration-dependent curves, with the highest background seen with the commercial clone MCA5748G. However, when using the Fab antibody molecule hFab as the standard (at the same range, from 1200 to 0.3 ng/mL), all the rabbit anti-hIgG clones again generated concentration-dependent curves. However, both the commercial mouse anti-hIgG clone MAC5748G and the control Gyrolab capture reagent, failed to produce a concentration-dependent curve.

Comparison of different clones used as capture reagents for detection of hIgG4 and hFab.
Dynamic range of rabbit anti-hIgG mAb clones in Gyrolab assay
One advantage of the Gyrolab assay is that it offers a significantly greater dynamic range when compared to the more common surface-based assay platforms (such as ELISA).(12) To assess the potential dynamic range of the clones, the human whole-antibody hIgG4 was used to prepare a linear 6-point curve 1:5 dilution with a range of 5000–0.32 ng/mL, as shown in Table 2. A similar Gyrolab assay procedure was performed using respective biotinylated capture antibody and the data were then extracted and compared. At the upper end (5000 ng/mL), the Gyrolab capture and the rabbit mAb clones 9E6, 2C5, and 19B1 had acceptable biases (<20% bias from nominal) and signal-to-noise ratios greater than 100. The three rabbit anti-hIgG clones showed better signal-to-noise ratios than the Gyrolab capture. At the 5000 ng/mL high endpoint, although clone 11G5 had an acceptable bias, its signal-to-noise ratio was low (63.3), which was caused by the relatively high signal level of the blank (7.4). For the same reason, commercial mAb clone MCA5748 also had a very poor signal-to-noise ratio at the upper end of the curve (24). These data suggest that clones 9E6, 2C5, and 19B1 offer a similar dynamic range when compared to the Gyrolab capture reagent, but with better signal-to-noise ratios. The dynamic range of detection for clones 9E6, 2C5, and 19B1 is significantly better compared with the commercial mouse anti-hIgG clone MCA5748G. In contrast, clones 16F5 and 11F9 performed well at the 0–1000 ng/mL range, but underrecovered at higher concentration levels. The differences in performance at higher concentration levels appear to correlate with epitope group, suggesting that dynamic range may be impacted by the antibody binding sites.
Comprising the Performance of Different Capture Antibodies by Gyrolab Assay
Values in italics indicate excellent performance, and values in bold indicate poor performance.
In conclusion, using the latest rabbit B cell cloning technology, we successfully developed six rabbit anti-hIgG mAb clones, which specifically recognize human IgG, but not cynomolgus monkey IgG. Amino acid sequence alignment demonstrates that all six clones are unique and diverse. Together, the six clones bind to three different epitope groups and are different from that of the commercial clone MCA5748G, suggesting that they may allow for development of pair-wise human IgG detection assays, such as “sandwich” ELISAs and Gyrolab assays. Initial evaluation of the performance of the clones using the Gyros platform indicates that they allow for specific and accurate detection of hIgG and hFab in a cynomolgus monkey serum matrix across a wide dynamic range. In contrast, the Gyros capture reagent and the commercial clone, MCA5748G, were only able to detect the whole human IgG, but not the Fab molecule, and S/N ratios suggest they have more cross-reactivity with monkey serum matrix. These data suggest that the newly developed antibody clones may provide a valuable resource in development of nonclinical drug concentration assays. With further assay development work, it is likely that sensitivity and dynamic range may be further optimized to align with expected drug concentration levels. Furthermore, it is expected that the newly developed rabbit anti-hIgG mAb clones can be used with matrix from other animal species.
Footnotes
Acknowledgments
We would like to send a warm thank you to Kathy Bankart for her ever valuable laboratory assistance. We would also like to extend our thanks to Andrew Xu for his valuable contributions and for his preliminary testing work.
Authors' Contributions
R.C. designed the project and performed some of the laboratory experiments. J.G. also performed some of the laboratory experiments. H.W., H.H., and Y.L. performed some of the laboratory experiments. R.P. supervised the project and provided management support.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.