
Abstract
Cytokeratin (CK) 18 is an intermediate filament protein that plays a major functional role in the integrity and mechanical stability of cells. Since both CK8 and CK18 are major components of simple epithelia, in the context of tumors, they are expressed in most carcinomas, and have been studied as diagnostic and prognostic markers in tumor pathology. CK18 is also cleaved by some caspases during apoptosis. Three-dimensional (3D)-cultured cancer cells are useful for cancer research as an intermediate model between in vitro cancer cell line cultures and in vivo tumors. In this study, we produced rat monoclonal antibodies (mAbs) through immunization of the lysate from 3D-cultured DLD-1 cells to elucidate a characteristic feature of a tumor, and our results showed that mAb 2H7 recognized human CK18. Furthermore, we indicated that mAb 2H7 was useful for immunoblotting, immunoprecipitation, and immunofluorescence staining. Therefore, it may be useful as a diagnostic tool for evaluating malignancy.
Introduction
Cytokeratins (CKs) are proteins found in the typical intermediate filament (IF) cytoskeletons of epithelial cells, which play a major functional role in cellular integrity and mechanical stability by attaching to nuclear membranes, epithelial cell–cell junctions, and desmosomes.(1,2) CKs also influence cell metabolic processes, cell growth, migration, and invasion.(2–4) CKs are classified as either acidic type I or basic or neutral type II, and they are heteropolymers by the association of type I and type II (1:1) molecules.(1) A pair of CKs, CK8 (type II, 52 kDa) and CK18 (type I, 45 kDa), are the major components of IFs of simple epithelia, and are not generally expressed in stratified epithelia. They are also expressed in mixed epithelia such as in the lung and breast, and are associated with cell differentiation. In tumors, CK8 and CK18 are expressed in most carcinomas such as adenocarcinomas, hepatocellular carcinomas, renal cell carcinomas, and neuroendocrine carcinomas.(1,5) Therefore, these CKs have been studied as diagnostic and prognostic markers in the context of tumor pathology.(1,6)
In cancer cells, CK18 is important for various cellular processes, such as apoptosis, mitosis, and cell cycle progression, through various cell signaling pathways, including the PI3K/Akt, Wnt, and ERK-MAPK signaling pathways.(7,8) Furthermore, CK18 is known to contribute to the initiation of transforming growth factor-β1–induced epithelial–mesenchymal transition in breast cancer.(9) During apoptosis, activated caspases are able to cleave CK18 at specific peptide recognition sites.(10,11) Caspase-cleaved CK18 is released into the circulation; therefore, it is useful to detect caspase-cleaved CK18 as a circulating biomarker of epithelium apoptosis. In a previous study, two sandwich enzyme-linked immunosorbent assays (ELISAs) were developed for detection of full-length and caspase-cleaved CK18.(12,13) Other studies have shown that the level of full-length and caspase-cleaved CK18 in the serum of patients was elevated in the presence of various cancers, and that this level can be considered a hallmark of malignancies.(14–17) Furthermore, increased serum caspase-cleaved CK18 has been shown to be a novel prognostic predictive biomarker after intracerebral hemorrhage, and an inflammatory marker of hepatitis B virus infection.(18,19) Thus, CK18 and its caspase-cleaved fragments have been reported as biomarkers for various diseases, and highly sensitive monoclonal antibodies (mAbs) against CK18 are available.
Human solid tumors develop and grow in three-dimensional (3D) microenvironments, such as tumor cells, nontumor cells, and extracellular matrix proteins. It has been reported to contain regions of low oxygen tension, termed as hypoxia.(20,21) Three-dimensional cell culture reflects many important properties of solid tumors, including the hypoxic gradient between the exterior and the center and the resistance against anticancer drugs and apoptosis.(22–25)
In this study, we generated a rat mAb 2H7 for response to human CK18 through immunization of the lysate from 3D-cultured DLD-1 cells as an intermediate model between in vitro cancer cell line cultures and in vivo tumors. Our results show that mAb 2H7 is useful for immunoblotting, immunoprecipitation (IP), and immunofluorescence staining. Therefore, we propose that this mAb can be useful for the diagnosis and the analysis of carcinogenesis in humans.
Materials and Methods
Cell culture
A172 (glioblastoma) and HepG2 (hepatoma) human cell lines were purchased from the RIKEN cell bank (Tsukuba, Japan). DLD-1 (colorectal adenocarcinoma), K562 (chronic myelogenous leukemia), and SUIT-2 (pancreatic carcinoma) human cell lines were purchased from the Cell Resource Center for Biomedical Research, Tohoku University (Sendai, Japan). LoVo (colorectal adenocarcinoma) human cell line was purchased from the Japanese Collection of Research Bioresources Cell Bank (Ibaraki, Japan). A172, DLD-1, and K562 cells were maintained in Roswell Park Memorial Institute (RPMI) 1640 medium (Fujifilm Wako, Osaka, Japan) and supplemented with 10% heat-inactivated fetal bovine serum (FBS, #SH30396; GE Healthcare, South Logan, UT) under standard culture conditions (5% CO2 at 37°C) until they reached confluence. HepG2, LoVo, and SUIT-2 cells were placed in Dulbecco's modified Eagle's medium (Fujifilm Wako) and cultured as already described.
Generation of rat mAb
Rat mAbs were generated using the rat lymph node method.(26,27) A cell membrane fraction of 3D-cultured DLD-1 cells was used as an antigen. Three-dimensional-cultured DLD-1 cells used to prepare the antigen lysate for antibody (Ab) production were maintained in RPMI medium with 10% FBS under standard culture conditions in a low-adsorption 24-well plate (PrimeSurface, Sumitomo Bakelite, Tokyo, Japan) for 6 days (12.8 × 104 cells/well). The cell membrane fraction was extracted according to the protocol from the subcellular protein fractionation kit for cultured cells (ThermoFisher Scientific, Eugene, OR).
An 8-week old female WKY/Izm rat (SLC, Shizuoka, Japan) was injected in the hind footpads with 3.5 mg of the cell membrane fraction of 3D-cultured DLD-1 cells. After 15 days, the cells from the medial iliac lymph nodes were fused with mouse myeloma SP2/0-Ag15 cells at a ratio of 5:1 with 50% polyethylene glycol (PEG1500; Roche, Indianapolis, IN) solution. The resulting hybridomas were plated onto 96-well plates (Corning, Kennebunk, ME) and cultured in HAT selection medium [Hybridoma-SFM (Life Technologies, Grand Island, NY), 10% FBS, BM Condimed H1 (Roche), Gibco HAT supplement (Life Technologies)]. At 8 days postfusion, the hybridoma supernatants were screened by immunoblotting against DLD-1 cells. As a result, mAb 2H7 was selected. The specific immunoglobulin class of mAb 2H7 was determined using a rat isotyping kit (BD Biosciences, San Diego, CA).
IP and identification of antigen
DLD-1 cells were seeded onto 10 cm dishes (one for each IP Ab) under standard culture conditions until they reached confluence, then lysed in 500 μL of lysis buffer [50 mM Tris, 150 mM NaCl, 1% NP-40, 0.5% sodium deoxycholate, and 0.1% sodium dodecyl sulfate (SDS) at pH 7.5] and inhibitor cocktail (Roche) on ice. Immunoprecipitations were performed with 40 μg of the rat mAb 2H7 or normal rat immunoglobulin G (IgG) (Fujifilm Wako) and 25 μL of the protein G-Sepharose beads (GE Healthcare). Proteins bound to beads were resolved by 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and then visualized by Coomassie brilliant blue (CBB) staining and immunoblotting. Desired bands were excised, in-gel-digested with trypsin (Promega, Madison, WI), and solvent extracted, and the resulting liquid chromatography–mass spectrometry (LC-MS) spectra obtained with an EASY-nLC 1000/Q-Exactive (ThermoFisher Scientific, Waltham, MA) were searched using the MASCOT search engine.
Immunofluorescence staining
DLD-1 cells were grown on a cell culture coverslip (Matsunami glass, Kishiwada, Japan) in RPMI medium with 10% FBS and fixed with 99.8% methanol for 5 minutes at −20°C. After blocking with phosphate-buffered saline containing 50 mM glycine, 1% BSA, and 2% normal goat serum, cells were incubated overnight with mAb 2H7 or anti-CK18 mouse mAb (1:1000 dilution, #NBP2-29461; Novus Biologicals, Centennial, CO) at 4°C.(28) Localization of antigens was observed using an Alexa488-conjugated goat antirat IgG (1:1000 dilution) or CF543-conjugated goat antimouse IgG (1:1000 dilution; Biotium, Hayward, CA). Immunofluorescence was detected using CKX53 fluorescent microscope (Olympus, Tokyo, Japan).
Immunoblotting
The sample was subjected to SDS-PAGE, using a 10% polyacrylamide gel, and subsequently transferred to Immobilon-P polyvinylidene fluoride transfer membrane (Millipore, Bedford, MA). The membrane was blocked overnight at 4°C with 3% skim milk in Tris-buffered saline with Tween-20 [TBS-T: 20 mM Tris-HCl (pH 7.5), 150 mM NaCl, 0.05% Tween-20], and incubated for 1 hour at room temperature with mAb 2H7 or anti-CK18 mouse mAb (1:500 dilution). After washing with TBS-T, the membranes were incubated for 30 minutes at room temperature with alkaline phosphatase (AP)-conjugated antirat IgG (1:5000 dilution; Sigma, St. Louis, MO) or AP-conjugated antimouse IgG (1:5000 dilution; Sigma). Membranes were then washed with TBS-T, and immunoreactivity was visualized with nitro blue tetrazolium chloride (Fujifilm Wako) and 5-bromo-4-chloro-3-indolyl phosphate (Fujifilm Wako) developing reagents.
Results and Discussion
To elucidate a characteristic feature of a tumor using 3D-cultured cancer cells, mAbs were produced against a lysate prepared from 3D-cultured DLD-1 cells, as described in Materials and Methods section. Because it is predicted to affect proteins located at the cell membrane or at the cytoskeletons of 3D-cultured cells compared with two-dimensional (2D)-cultured cells, we selected the membrane fraction of a lysate prepared from 3D-cultured cells for our antigen. Fifteen days postimmunization, lymphocytes were collected from the enlarged lymph nodes of the rat, then hybridomas were obtained after fusing the lymphocytes with mouse myeloma SP2/0-Ag15 cells. These obtained hybridomas were evaluated to see whether they produced mAbs that recognized DLD-1 cells by immunoblotting. The Ab, designated mAb 2H7, gave a signal corresponding to ∼45 kDa and its specific immunoglobulin class was rat IgG1 (κ) class.
To identify the antigen recognized by mAb 2H7, we performed an IP of whole-cell lysate prepared from DLD-1 cells with mAb 2H7. The protein that interacted with mAb 2H7 was detected by CBB staining. As a result, a protein with molecular mass of 45 kDa was detected (Fig. 1A left panel). Immunoblotting indicated that the antigen was recognized by mAb 2H7 (Fig. 1A right panel). The relevant band was excised, analyzed by LC-MS, and identified as a CK18. Next, we confirmed whether the antigen of mAb 2H7 was in fact CK18. As already mentioned, IP was performed using the DLD-1 lysate for both mAb 2H7 and purchased anti-CK18 mouse mAb, followed by immunoblotting for both mAbs. This result confirmed that a band with molecular mass of 45 kDa was a CK18 (Fig. 1B). We performed a series of these experiments testing rat IgG as a negative control Ab, in which no signal was obtained from the IP (Fig. 1A, B).
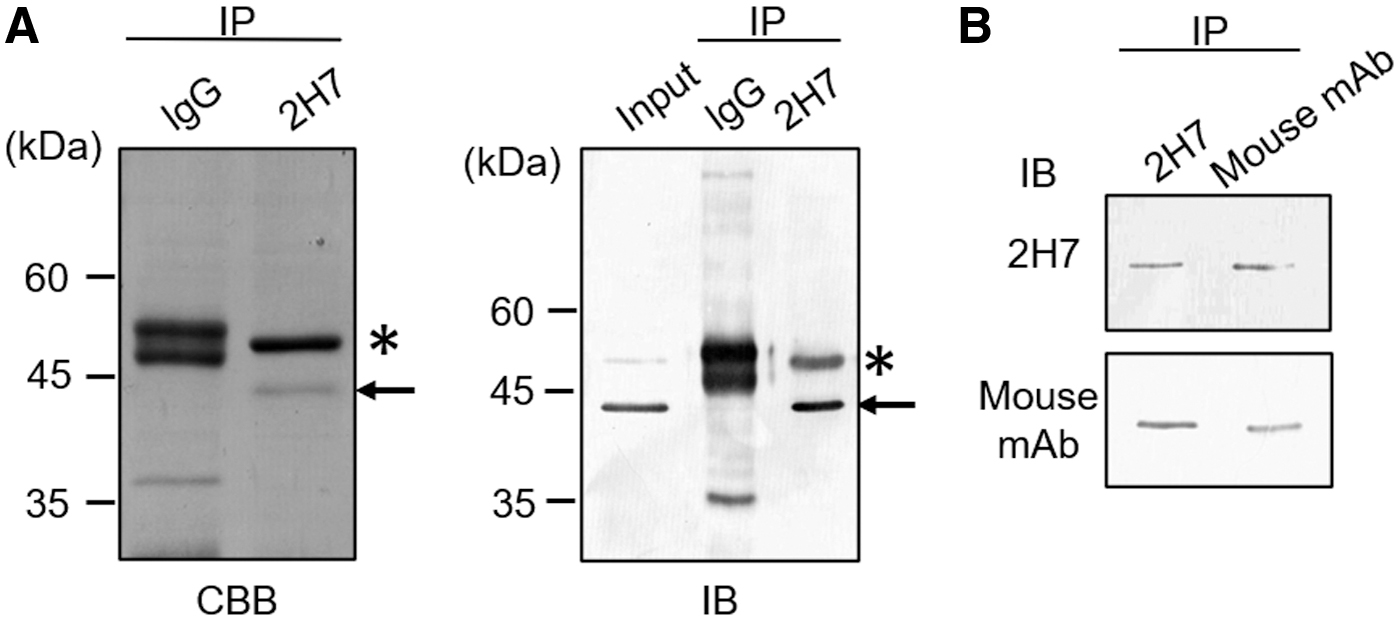
Identification of antigen recognition by mAb 2H7.
Next, we examined whether mAb 2H7 reacts with endogenous CK18, using whole-cell lysates from the various cell lines. Whole-cell lysates were prepared from A172, DLD-1, HepG2, K562, LoVo, and SUIT-2 cells. As shown in Figure 2A, mAb 2H7 exclusively recognized CK18, which was present in all lysates, as well as the mouse mAb. These results show that mAb 2H7 recognized endogenous CK18 in these cell lines. Furthermore, this experiment suggested that mAb 2H7 could also recognize CK8, as indicated by the band with 50 kDa, particularly in DLD-1 cells. Owing to the high homology of CK8 and CK18 in humans, some Abs raised against CK18 cross-reacted with CK8.(29) Furthermore, mAb 2H7 recognized a band with molecular mass of ∼43 kDa in DLD-1, HepG2, LoVo, and SUIT-2 cells. Since it is known that CK18 is cleaved by activated caspases during apoptosis, we assume that a band with molecular mass of ∼43 kDa is a cleaved product, and because these are induced during apoptosis, they can be markers of apoptosis. In addition, these products have been investigated and are related to poor prognoses. Finally, immunofluorescence staining was performed to confirm the localization of CK18 in DLD-1 cells. Our results showed that mAb 2H7 stained the cytoplasm in a mesh pattern, and that mouse mAb showed a similar staining pattern (Fig. 3). Thus, we propose in conclusion that the antigen of mAb 2H7 is CK18.
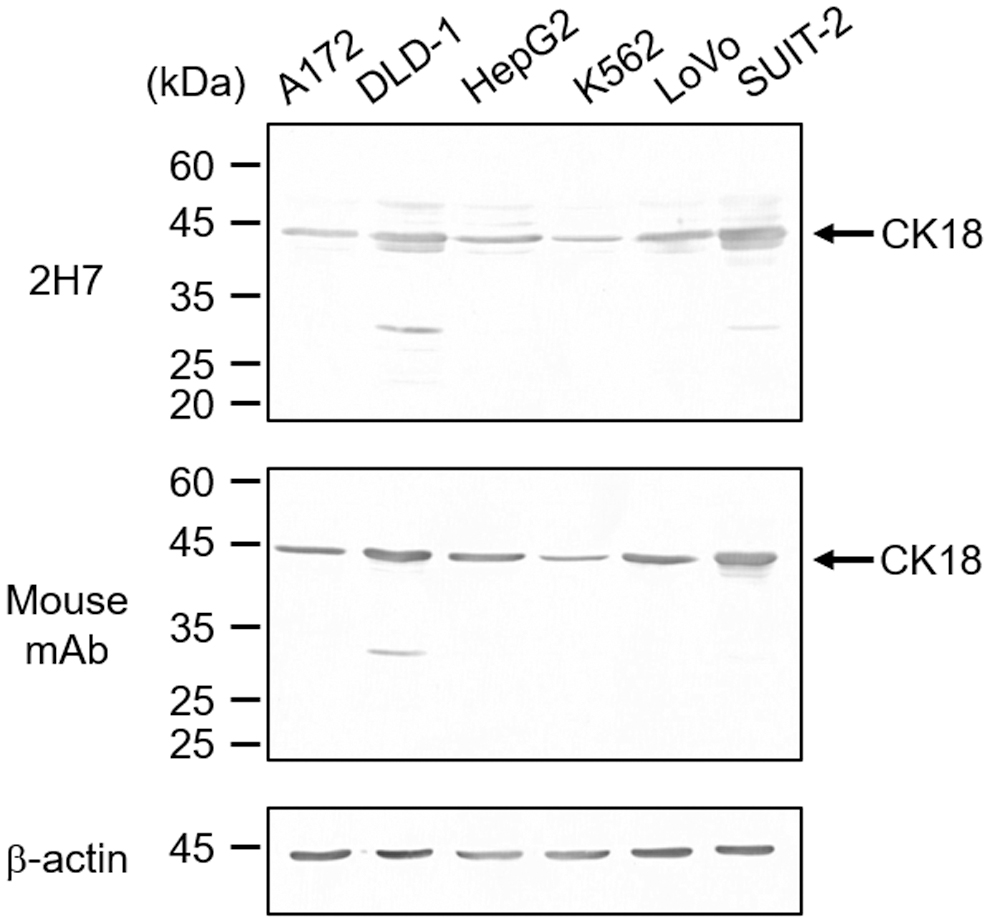
Comparison of recognition of endogenous CK18 by mAb 2H7 and mouse mAb through immunoblotting using several human cell lines. Lysates prepared from A172, DLD-1, HepG2, K562, LoVo, and SUIT-2 cells were separated on 10% SDS-PAGE gel and immunoblotted with mAb 2H7, anti-CK18, and anti-b-actin mAbs. Locations of the molecular weight markers are indicated at the left of the panels.
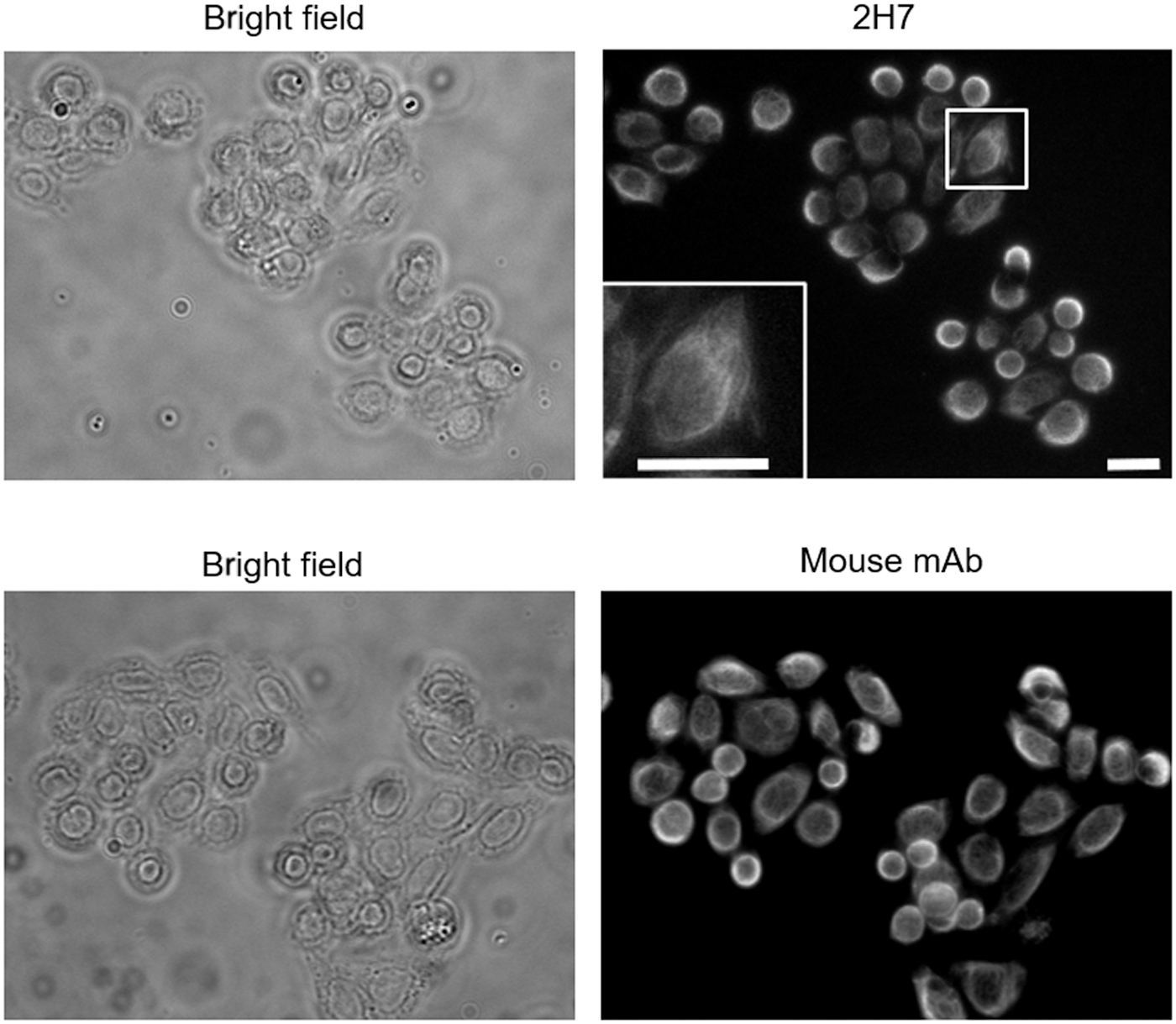
Immunofluorescence staining of mAb 2H7 and mouse mAb using DLD-1 cells. DLD-1 cells were immunostained with mAb 2H7 or anti-CK18 mAb. The rat and mouse anti-CK18 mAbs were detected using Alexa488-conjugated antirat IgG and CF543-conjugated antimouse IgG, respectively. Inset is the magnification of the white square. Scale bar: 20 μm.
In summary, we generated rat mAb 2H7, which recognized CK18 by immunization of 3D-cultured cancer cells. CK18 is known to relate to carcinogenesis, through various regulating signals. Therefore, we propose that mAb 2H7 could be generated by the immunization of 3D-cultured cancer cells as a tumor model in vitro. Furthermore, gene expression profiles of the tumor obviously differed from conventional 2D cultures but were similar to 3D cultures.(30,31) However, it remains unclear about the expression levels of CK18 in 2D- and 3D-cultured cancer cells. In future studies, it is necessary to perform quantitative PCR on CK18 mRNA and on mRNAs of other CKs in 3D-cultured cancer cells compared with 2D-cultured cancer cells. We also observed that mAb 2H7 may not recognize mouse, rat, dog, and monkey CK18 (data not shown). CK18 and its caspase-cleaved fragments have been reported as biomarkers for various diseases including cancer malignancy, and diagnoses using anti-CK18 Abs are available. Yoon et al. indicated the significant expression of CK18 could provide useful markers for differentiating ameloblastic carcinoma using anti-CK18 mouse mAb purchased from Novus Biologies.(28) We believe that mAb 2H7 should prove useful for the diagnosis and the analysis of cancer in humans because mAb 2H7 reacts as well as commercial anti-CK18 mouse mAb.(28)
Footnotes
Authors' Contributions
K.S. carried out data curation, formal analysis, methodology, and writing—review and editing—of the article. K.I. was involved in writing—review and editing—of the article. H.H. conceptualized and wrote—review and editing—the article. C.Y. performed conceptualization, data curation, formal analysis, funding acquisition, methodology, supervision, project administration, and writing—original draft, and review and editing—of the article.
Author Disclosure Statement
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this article.
Funding Information
This study was supported by Grants 20K07583 from the Japan Society for the Promotion of Science KAKENHI (C.Y.), the Sasakawa Scientific Research Grant from the Japan Science Society (C.Y.), and the Chiome Grant from LNest (C.Y.).