
Abstract
Immunoglobulin A (IgA) antibodies are critical to mucosal protection, specifically dimeric IgA (dIgA) and secretory IgA (sIgA), which rely on the J chain to polymerize. There is an absence of monoclonal antibodies that can specifically bind to polymeric IgA without the need to denature the molecule. We generated a panel of highly specific mouse anti-J chain antibodies that react with both intact and denatured nonhuman primate dIgA and human dIgA and sIgA of both the IgA1 and IgA2 subclass. We expanded use of this antibody for quantification of dIgA and sIgA using biolayer interferometry or enzyme-linked immunosorbent assay and use for affinity chromatography. This is a significant improvement over available anti-IgA antibodies in the field, which will allow for expanded use in clinical testing.
Introduction
The Immunoglobulin A (IgA) isotype of immunoglobulins plays a critical role in mucosal immunity and immune responses.(1,2) In addition to monomeric molecules, IgA can form polymeric molecules such as dimers and tetramers by assembling with a 15 kD polypeptide referred to as Joining (J) shortly before or at the time of secretion by the antibody-producing cell [reviewed in Woof and Mestecky(2)]. Dimeric IgA (dIgA) binds to the polymeric immunoglobulin receptor (pIgR) on the basolateral surface of epithelial cells. Once bound, the dIgA is transported through the cell and released on the apical (mucosal) side, along with the cleaved portion of the pIgR referred to as secretory component (SC, 70–80 kD).(3) Although SC is not physically associated with the J chain of the dIgA, J chain is necessary for secretory IgA (sIgA) formation. sIgA is involved in the adaptive (antigen binding) and innate (adhesion due to glycans) arms of the immune system.(4) Several polyclonal and monoclonal antibody preparations available commercially react with the J chain; however, with the exception of one published(5) or commercial antibody, they only react with denatured IgA.
In this study, we report a monoclonal antibody that specifically reacts with the J chain of intact native dIgA and sIgA, in addition to denatured molecules. Furthermore, this antibody reacts with dIgA and sIgA of both subclasses (IgA1 and IgA2) as well as nonhuman primate J chain. We demonstrate that this antibody can be used not only for Western blot but also to specifically quantify and detect dIgA and sIgA by enzyme-linked immunosorbent assay (ELISA) or biolayer interferometry (BLI). It can also be used for the purification of dIgA from a mixed population. This is a significant improvement on current commercial products that fail to react with native J chain limiting their utility.(6)
Materials and Methods
Expression and purification of J chain fusion proteins from bacteria
The nucleic acid sequences of J chain from (NP_690052.2) were synthesized (Genscript) and cloned into a pMAL-c5x vector (Addgene) in-frame with a maltose binding protein (MBP). The cloned J chain construct was transformed into ShuffleT7 competent Escherichia coli cells (New England Biolabs [NEB]) and expression was induced with 0.1 mM isopropyl β-D-1-thiogalactopyranoside. Bacteria were lysed and proteins were incubated with amylose resin (NEB) for 2 hours. Purified J chain was then eluted with 20 and 50 mM maltose elution buffer (Sigma). Proteins were dialyzed in phosphate-buffered saline (PBS) for 2 hours at room temperature then stored in −20°C.
Mouse immunization
CD1 mice (Charles River) were immunized with 50 μg of J chain MBP with the Sigma adjuvant system weekly for 5 weeks. The anti-J chain titer in mouse serum was measured by ELISA on week 6. A final intraperitoneal/intramuscular injection was administered 48 hours before fusion. Splenocytes from mice with the highest serum titer to J chain MBP were fused with P3X partner cells. Hybridomas were generated following a standard polyethylene glycol fusion protocol.(7) Two weeks postfusion, hybridomas were screened for reactivity to J chain as well as MBP. Positive cell clones against J chain were selected for antibody sequencing and further testing against polymeric forms of IgA.
Enzyme-linked immunosorbent assay
Ninety-six-well plates (Nunc) were coated with 2 μg/mL of purified J chain MBP or MBP only and incubated at 4°C overnight. The plates were blocked with 1% bovine serum albumin plus 0.05% Tween20 in PBS. Hybridoma supernatants were added neat to the plates, whereas purified antibodies were diluted in PBS plus 0.1% Tween20 before being added. Samples were incubated for 1 hour at room temperature. The plates were stained with horseradish peroxidase (HRP) goat antimouse IgG Fcγ (Southern Biotechnology) for 1 hour and developed using 3,3′,5,5′-tetramethylbenzidine substrate (ThermoFisher). Absorbance at 450 nm (optical density450) was measured on an Emax precision plate reader (Molecular Devices). Hybridomas that bound MBP J chain, but not MBP only, were selected. Antibody isotype was determined using Pierce™ Rapid Antibody Isotyping Kit per the manufacturers' instruction.
Antibody cloning and expression
Initial selection of positive hybridomas and clones was performed with antibodies expressed by hybridoma. After confirmation of positive clones, the heavy chain and light chain variable regions were amplified from hybridoma cells and cloned into two pcDNA 3.1 (ThermoFisher) vectors containing κ light chain constant region and IgG1 heavy chain constant region, respectively, as previously described.(7) Antibody was expressed by cotransfecting human kidney 293 cells (HEK293) using stable cationic polymer(8) polyethyleneimine (Sigma-Aldrich). Supernatants of transfected cells were tested for reactivity and harvested 7 days post-transfection. Antibody was purified from the supernatants using protein G sepharose beads (GE Healthcare). Beads were washed with PBS to remove nonspecific proteins and antibodies were eluted with 0.1 M glycine pH 2.8.
Immunoblotting
IgA proteins (5 μg) were either not reduced or reduced using 5% β-mercaptoethanol followed by heating at 95°C for 5 minutes. Samples were then loaded into NuPAGE 12%, Bis–Tris, 1.0 mm, Mini Protein Gel (Invitrogen NP0341PK2). Samples were transferred and then blocked in PBS with 5% milk for 20 minutes then stained with the purified mouse anti-J chain antibody CA1L33e1 for 1 hour. Bound anti-J chain antibody was detected with antimouse HRP (cat. no. 1030-05; Southern Biotech) at 1:5000 dilution conjugated, then the image was taken after developing with ECL substrates (cat. no. A38554; Bio-Rad).
Biolayer interferometry
BLI with an Octet HTX (PALL/ForteBio) was used to determine the concentration of dIgA forms. Anti-J chain antibody was diluted into PBS at 20 μg/mL and captured to an antimouse IgG Fc capture (AMC) biosensors (cat. no. 18-5088; ForteBio) that were dipped to a range of known dIgA concentrations (3–100 μg/mL) to generate a standard curve. This standard curve was then used to quantify an unknown range of dIgA concentrations.
Statistical analysis
Statistical analyses were performed using GraphPad Prism v8.1.1. Data are plotted as mean ± standard deviation of n = 3 independent runs where applicable.
Results and Discussion
An immunogen was developed by expressing the human J chain coding region in a pMAL-5 vector containing an MBP tag. The J chain MBP product was purified using an amylose resin, quantitated by ultraviolet-visible spectroscopy, mixed with Ribi adjuvant, and subsequently used to immunize mice. Mice were initially immunized by intraperitoneal injection (IP) with 50 μg/mL J chain MBP fusion followed by weekly boosts of 25 μg/mL for 3 weeks. Mice were boosted with J chain MBP by intravenous injection and IP 3 and 2 days before fusion. Sera from the immunized mice demonstrated a robust response against J chain MBP (Fig. 1A, B) as compared with sera from unimmunized mice.
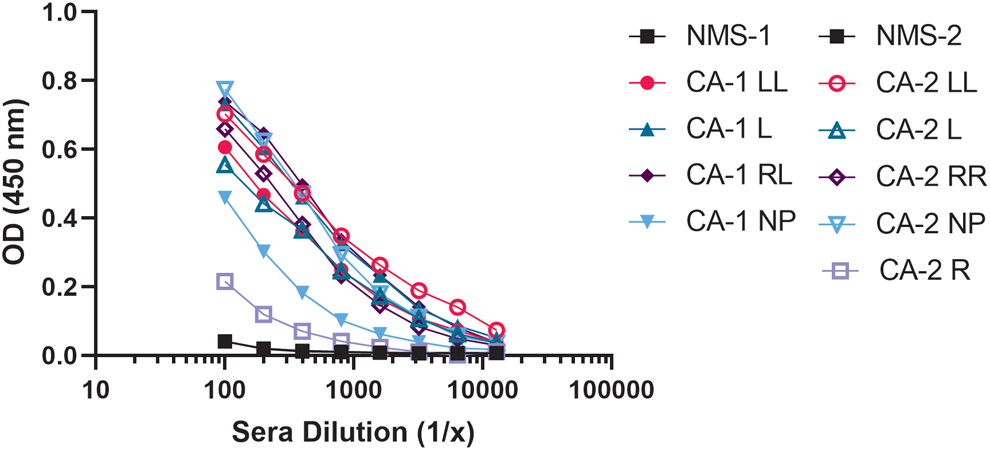
Reactivity of sera from immunized mice with J chain. Eleven CD1 mice were immunized by intraperitoneal injection with 50 μg of J chain MBP fusion in Ribi adjuvant. After 6 weeks, sera were collected and tested by ELISA for presence of antigen-specific antibody. Plates were coated with purified J chain protein and bound antibody was detected using goat antimouse IgG-HRP conjugate. Immunized mice (CA1 and CA2) developed higher IgG serum titers against J chain protein than unimmunized control mice (NMS, black). Mice were labeled based on CA and ear piercing. Data are plotted as mean of n = 3 independent runs. The CV was <10% for the samples. CA, cage; CV, coefficient of variation; ELISA, enzyme-linked immunosorbent assay; HRP, horseradish peroxidase; IgG, immunoglobulin G; L, left ear; LL, double tags on left ear; MBP, maltose binding protein; NMS, normal mouse serum; NP, no tags; OD, optical density; R, right ear; RR, double tags on right ear.
Four separate fusions were performed with four mice (CA1L, CA1RL, CA1LL, and CA1NP) using a 3:1 ratio of splenocytes to fusion partner (P3X) resulting in 40 plates per fusion. Supernatants from the hybridomas were screened for reactivity against J chain and dIgA. The first round of antibody screening was to select for antibodies specific to J chain only. Further selection was based on reactivity with intact J chain in dIgA1 and dIgA2 subclass of at least two different human monoclonal IgA antibodies. Lead hybridomas were tested further for reactivity with dIgA1, dIgA2, rhesus dIgA, and against monomeric IgA (mIgA) to ensure specificity. Selected antibodies failed to react with mIgA. As shown in Figure 2, purified murine antibodies CA1L33e1 and CA1LL23h8 as well as positive control serum from CA1LL mouse reacted with both subclasses of dIgA, with CA1L33e1 antibody showing higher binding of the two murine antibodies. Therefore, clone CA1L33e1 was selected as the lead anti-J chain antibody. Limiting dilutions of the CA1L33e1 antibody were used to isolate a high producing homogeneous clone.
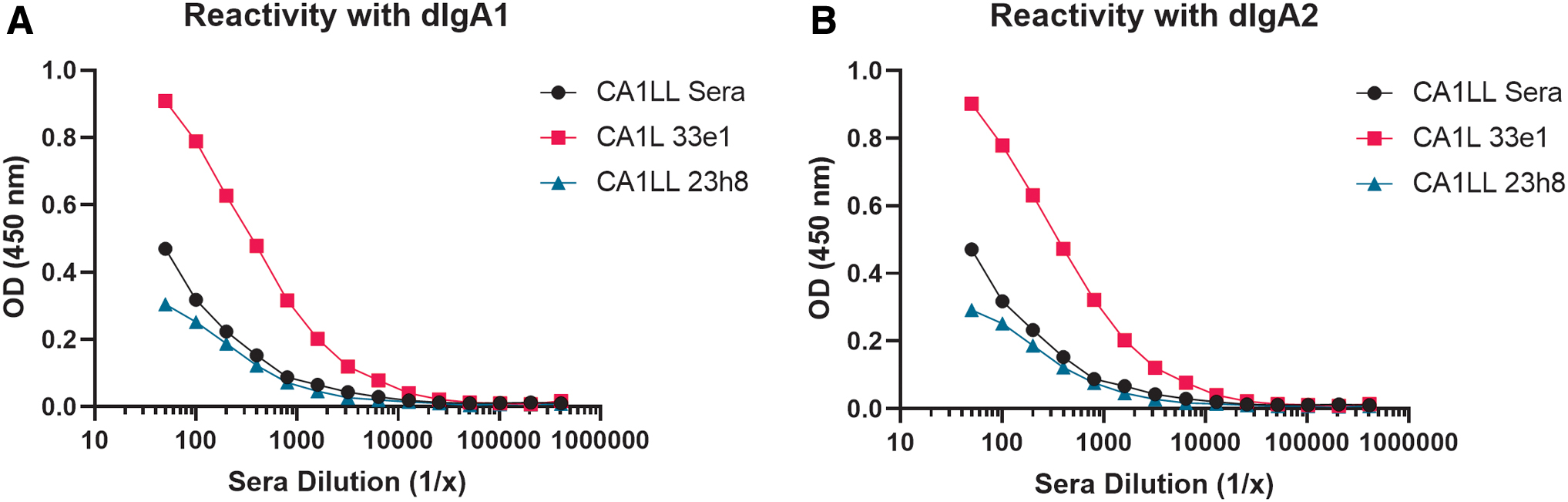
Anti-J chain antibody reactivity with human IgA subclasses. Lead purified anti-J chain antibodies CA1L clone 33e1 and CA1LL clone 23h8 were tittered against dIgA1
There is 98% homology between rhesus J chain and human J chain (Fig. 3A); therefore, it is not a surprise that mouse anti-J chain CA1L33e1 also reacts with rhesus dIgA (Fig. 3B). This antibody can be used for similar applications in rhesus as well. Furthermore, in addition to reacting with dIgA of both subclasses, CA1L33e1 also reacts with sIgA of both subclasses (Fig. 4). The lack of reagents for detection of native dIgA or sIgA has been a challenge for high throughput or standardized measurement of dIgA or sIgA in biological samples or during the antibody purification process. Therefore, we tested the CA1L33e1 antibody for use as a detecting antibody for immunoblot, ELISA, BLI (Octet), and affinity chromatography.

Reactivity of anti-J chain with J chain of nonhuman primates.

Reactivity of CAIl33e with sIgA. ELISA plates were coated with mIgA, dIgA, and sIgA followed by addition of anti-J chain mouse antibody CA1L33e1. Bound antibody was detected as shown in Figure 1. Data are plotted as mean ± standard deviation of n = 3 independent runs.
To determine the utility of CA1L33e1 for immunoblots, mIgA, dIgA, and sIgA from both IgA1 and IgA2 subclasses were subjected to electrophoresis under nonreducing (Fig. 5A) and reducing (Fig. 5B) conditions. After transfer, the blots were incubated with CA1L33e1 followed by goat antimouse IgG HRP. As shown in Figure 5, CA1L33e1 reacted with the J chain in dimeric and secretory forms of human IgA1 and IgA2 under both conditions. This is in contrast to several commercial antibodies that only react under reducing conditions. As expected, there was no reactivity with mIgA. Finally, consistent with ELISA results, CA1L33e1 reacted with rhesus dIgA but not with mIgA.
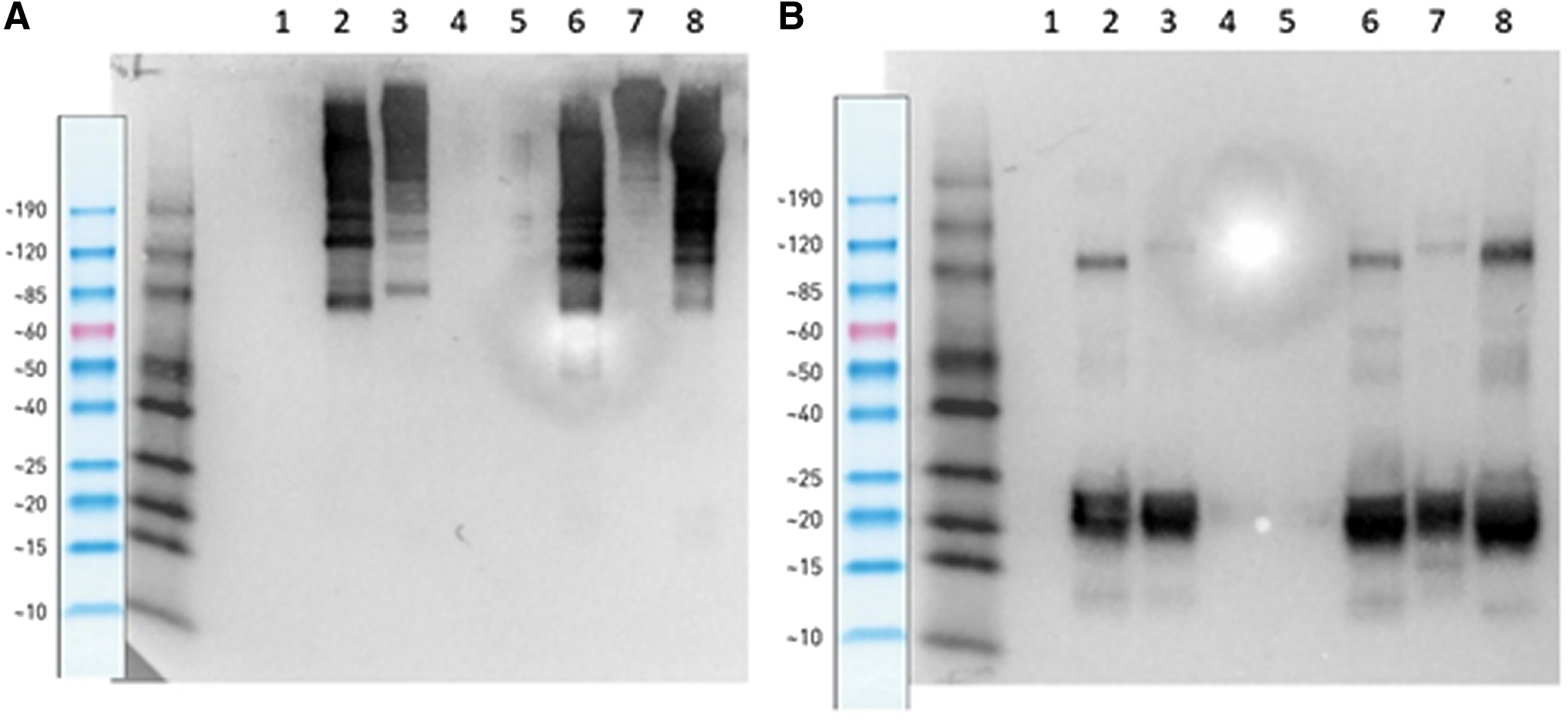
Immunoblot of anti-J chain antibody CA1L33e1 with dimeric and secretory IgA1 and IgA2. Immunoblot of human IgA2 (lanes 5–7) and IgA1 (lanes 1–3), in monomeric (lanes 1 and 5), dimeric (lanes 2 and 6), and secretory (lanes 3 and 7) form on SDS-PAGE under nonreducing
Known concentrations of mIgA and dIgA were combined to determine the sensitivity of detection of dIgA by ELISA. As shown in Figure 6, CA1L33e1 accurately measured the amount of dIgA in the sample, regardless of the presence of mIgA. Sensitivity was >0.1 μg/mL. We have successfully used this antibody to measure the levels of dIgA in both rhesus and human samples (plasma and mucosal secretions) as well as to selectively quantitate the dIgA content of antibody preparations by ELISA (data not shown). Similarly, a standard curve using known concentrations of dIgA was generated using the Octet and accurately determined the concentration of spiked dilutions of dIgA (data not shown).

Use of anti-J chain antibody for dIgA quantitation. ELISA plates were coated with antigen (LT) before the addition of LT-specific antibody. Test antibody consisted of either monomer or dimer alone or a combination of the two. Bound antibody was detected using CA1L33e1 and ELISA continued as shown in Figure 1. Data are plotted as mean ± standard deviation of n = 3 independent runs. LT, heat labile toxin.
CA1L33e1 can also be used to purify dIgA from these same sources after coupling to an immobilized bead such as sepharose. For some applications, it may be beneficial to use a smaller active fragment of the antibody. The Fab fragment of CA1L33e1 captures 70% of the antibody compared with IgG. Both the Fab and the IgG of CA1L33e1 were successfully coupled to sepharose and used to purify dIgA from supernatant (data not shown). Thus, the Fab fragment represents the best choice for a smaller antibody format for affinity matrix applications.
Summary
CA-1L 33e A1a3 antibody has shown many advantages over any commercially available antibodies against J chain. Its specificity enables it to be used in a variety of different ways. It is a unique antibody that can be used to quantify and detect the presence of both subclasses and forms of IgA in mucosal samples from clinical trials as well as the measurement of antigen-specific B cell response.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the Defense Advanced Research Project Agency (Grant No. DARPA-BAA-13-03) and the Bill & Melinda Gates Foundation (Grant No. OPP1173647).