
Abstract
CC chemokine receptor 9 (CCR9) belongs to the beta chemokine receptor family and is mainly distributed on the surface of immature T lymphocytes and enterocytes. This receptor is highly expressed in rheumatoid arthritis, colitis, type 2 diabetes, and various tumors. Therefore, more sensitive monoclonal antibodies (mAbs) need to be developed to predict the prognosis of many high CCR9 expression diseases. Because CCR9 is a structurally unstable G protein-coupled receptor, it has been difficult to develop anti-CCR9 mAbs using the traditional method. This study developed anti-human CCR9 (hCCR9) mAbs for flow cytometry using a Cell-Based Immunization and Screening (CBIS) method. Two mice were immunized with hCCR9-overexpressed Chinese hamster ovary (CHO)-K1 cells (CHO/hCCR9), and hybridomas showing strong signals from CHO/hCCR9 and no signals from CHO-K1 cells were selected by flow cytometry. We established an anti-hCCR9 mAb, C9Mab-1 (IgG1, kappa), which detected hCCR9 in MOLT-4 leukemia T lymphoblast cells and CHO/hCCR9 cells by flow cytometry. Our study showed that an anti-hCCR9 mAb was developed more rapidly by the CBIS method than the previous method.
Introduction
Chemokines are classified into four groups, CXC, CC, C, and CX3C chemokines, and are responsible for regulating cellular trafficking of various types of leukocytes.(1–4) Among them, CC chemokines are composed of 27 chemotactic cytokines(3,4) and have a C-C structure in which the first two cysteines are adjacent to each other.(5) CC chemokine receptor 9 (CCR9) is a G protein-coupled receptor (GPCR) distributed mainly on the surface of immature T lymphocytes and enterocytes. It belongs to the beta chemokine receptor family.(6) CC chemokine ligand 25 (CCL25), a member of the CC chemokine subfamily, is also known as a thymus-expressed chemokine and is the only ligand for CCR9.(7) CCR9 and CCL25 play essential roles in inflammatory cell proliferation and chemotaxis.(8)
Overexpression of CCR9 and CCL25 is observed in many cancers, including leukemia,(9) melanoma,(10) prostate cancer,(11) breast cancer,(12) lung cancer,(13) ovarian cancer,(14) and hepatocellular carcinoma.(12) CCR9 is also expressed in cells of various other tissues, such as the trophoblast,(15) heart,(16) small intestine,(17) and endometrial stroma,(18) and promotes cell proliferation,(19–22) tumorigenesis,(13,19,22) metastasis,(11,14,19,23,24) and chemoresistance in tumors.(25–28) Furthermore, CCR9 is useful for predicting lymph node metastasis in lung adenocarcinoma and may serve as a new prognostic biomarker.(13)
Also, CCR9 is highly expressed in rheumatoid arthritis, colitis, and type 2 diabetes. In vivo studies have shown that it may be an effective therapeutic target.(29–32) Therefore, CCR9 may be useful as a diagnostic agent to predict the prognosis of many diseases, and more sensitive monoclonal antibodies (mAbs) need to be developed. In this study, we established a sensitive and specific mAb against human CCR9 (hCCR9).
Materials and Methods
Cell lines
Chinese hamster ovary (CHO)-K1 and P3X63Ag8U.1 (P3U1) were obtained from the American Type Culture Collection (ATCC, Manassas, VA). MOLT-4 (a leukemia T lymphoblast cell line) was obtained from the Japanese Collection of Research Bioresources (JCRB) Cell Bank (Osaka, Japan). The expression plasmid of hCCR9 (pCMV6neoCCR9-Myc-DDK) was purchased from OriGene Technologies, Inc., (Rockville, MD) and was transfected using Lipofectamine LTX with Plus Reagent (Thermo Fisher Scientific, Inc., Waltham, MA). Stable transfectants were selected by limiting dilution and cultivation in a medium containing 0.5 mg/mL of G418 (Nacalai Tesque, Inc., Kyoto, Japan).
CHO-K1, P3U1, CHO/hCCR9, and MOLT-4 were cultured in Roswell Park Memorial Institute (RPMI) 1640 medium (Nacalai Tesque, Inc.), supplemented with 10% heat-inactivated fetal bovine serum (Thermo Fisher Scientific, Inc.), 100 U/mL penicillin, 100 μg/mL streptomycin, and 0.25 μg/mL amphotericin B (Nacalai Tesque, Inc.). Cells were grown in a humidified incubator at 37°C with an atmosphere of 5% CO2 and 95% air.
Antibodies
Anti-hCCR9 mAb (clone #112509) was purchased from R&D systems (Minneapolis, MN). Secondary Alexa Fluor 488-conjugated anti-mouse IgG was purchased from Cell Signaling Technology, Inc. (Danvers, MA).
Hybridoma production
Female BALB/c mice (6 weeks old) were purchased from CLEA Japan (Tokyo, Japan). The animals were housed under specific pathogen-free conditions. All animal experiments were performed according to relevant guidelines and regulations to minimize animal suffering and distress in the laboratory. Animal experiments were approved by the Animal Care and Use Committee of Tohoku University (Permit number: 2019NiA-001). Mice were monitored daily for health during the full 4-week duration of the experiment. A reduction of more than 25% of total body weight was defined as a humane endpoint. Mice were euthanized by cervical dislocation, and death was verified by respiratory and cardiac arrest.
To develop mAbs against hCCR9, we employed the Cell-Based Immunization and Screening (CBIS) method.(33) Briefly, two BALB/c mice were immunized with CHO/hCCR9 cells (1 × 108) by the intraperitoneal route together with the Imject Alum (Thermo Fisher Scientific, Inc.). The procedure included three additional immunization followed by a final booster intraperitoneal injection administered 2 days before spleen cells were harvested. Harvested spleen cells were subsequently fused with P3U1 cells using PEG1500 (Roche Diagnostics, Indianapolis, IN). The hybridomas were grown in an RPMI medium supplemented with hypoxanthine, aminopterin, and thymidine for selection (Thermo Fisher Scientific, Inc.). Culture supernatants were screened by flow cytometry.
Flow cytometry
Cells were harvested after brief exposure to 0.25% trypsin and 1 mM ethylenediaminetetraacetic acid (Nacalai Tesque, Inc.). They were washed with 0.1% bovine serum albumin in phosphate-buffered saline and treated with primary mAbs for 30 minutes at 4°C. The cells were then treated with Alexa Fluor 488-conjugated anti-mouse IgG (1:2000; Cell Signaling Technology, Inc.). Fluorescence data were collected using the SA3800 Cell Analyzer (Sony Corp., Tokyo, Japan).
Determination of binding affinity by flow cytometry
CHO/hCCR9 was suspended in 100 μL of serially diluted anti-hCCR9 mAbs (0.006–50 μg/mL), and Alexa Fluor 488-conjugated anti-mouse IgG (1:200; Cell Signaling Technology, Inc.) was added. Fluorescence data were collected using the BD FACSLyric (Becton, Dickinson and Company, Franklin Lakes, NJ, USA). The dissociation constant (KD) was calculated by fitting binding isotherms to built-in, one-site binding models in GraphPad PRISM 8 (GraphPad Software, Inc., La Jolla, CA).
Results
Establishment of anti-hCCR9 mAbs
To produce anti-hCCR9 mAbs, we employed the CBIS method using stable transfectants for both immunization and flow cytometric screening (Fig. 1). Two mice were immunized with CHO/hCCR9 cells, which overexpress hCCR9. Hybridomas were seeded into 96-well plates, and CHO/hCCR9-positive and CHO-K1-negative wells were selected. After limiting dilution, C9Mab-1 (IgG1, kappa) was finally established.
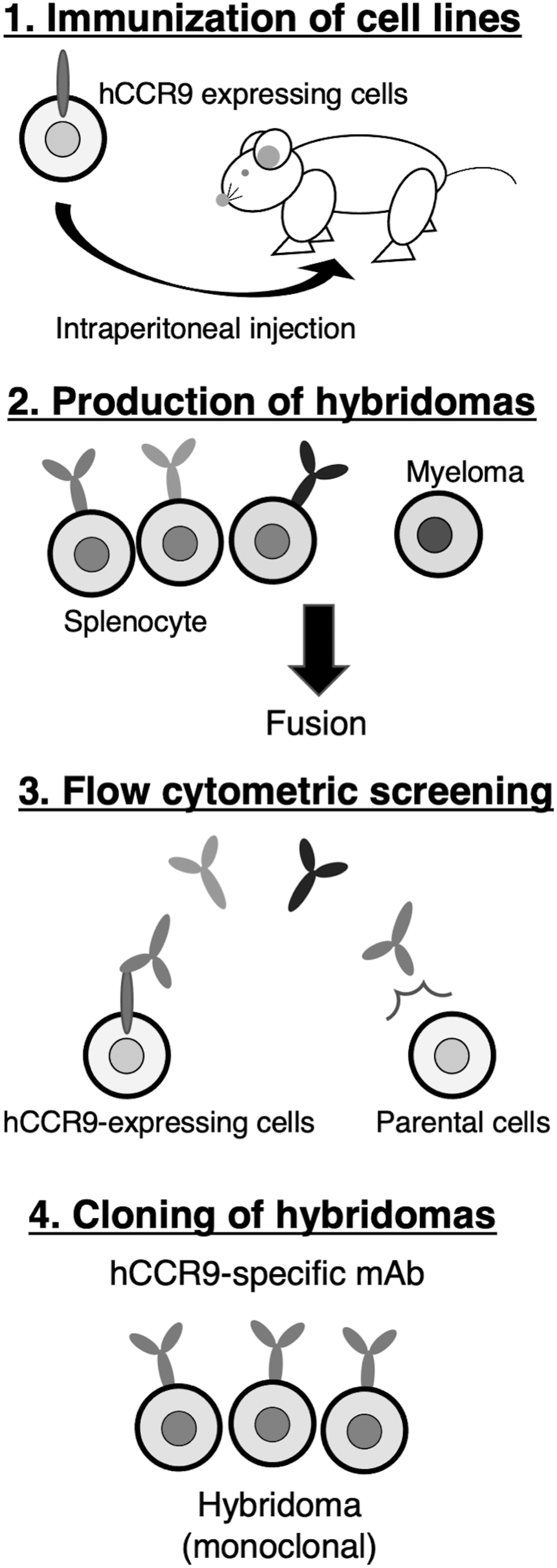
Production of anti-hCCR9 mAbs. The procedure of the CBIS method. CHO/hCCR9 cells were immunized into BALB/c mice using intraperitoneal injection. The screening was performed by flow cytometry. CBIS, Cell-Based Immunization and Screening; CHO, Chinese hamster ovary; hCCR9, human CC chemokine receptor 9; mAbs, monoclonal antibodies.
Flow cytometry
We performed flow cytometry using C9Mab-1 against CHO/hCCR9, CHO-K1, and MOLT-4. C9Mab-1 recognized CHO/hCCR9 dose-dependently (Fig. 2A), but not CHO-K1 (Fig. 2B). Another anti-hCCR9 mAb (clone #112509 from R&D systems: positive control) also recognized CHO/hCCR9 dose-dependently (Fig. 2A), but not CHO-K1 (Fig. 2B). Although C9Mab-1 reacted with CHO/hCCR9 even at 0.001 μg/mL, clone #112509 reacted with it in more than 0.1 μg/mL (Fig. 2A), indicating that the sensitivity of C9Mab-1 is higher than that of clone #112509. C9Mab-1 also reacted with MOLT-4 cells, which express endogenous hCCR9 dose-dependently even in 0.001 μg/mL (Fig. 2C). In contrast, clone #112509 reacted with it in more than 0.1 μg/mL (Fig. 2C).

Flow cytometry using anti-hCCR9.
Determination of the binding affinity of C9Mab-1
We then assessed the apparent binding affinity of C9Mab-1 with CHO/hCCR9 using flow cytometry. The KD of C9Mab-1 for CHO/hCCR9 was 1.2 × 10−9 M (Fig. 3), indicating that C9Mab-1 possesses a high affinity for CHO/hCCR9 cells.
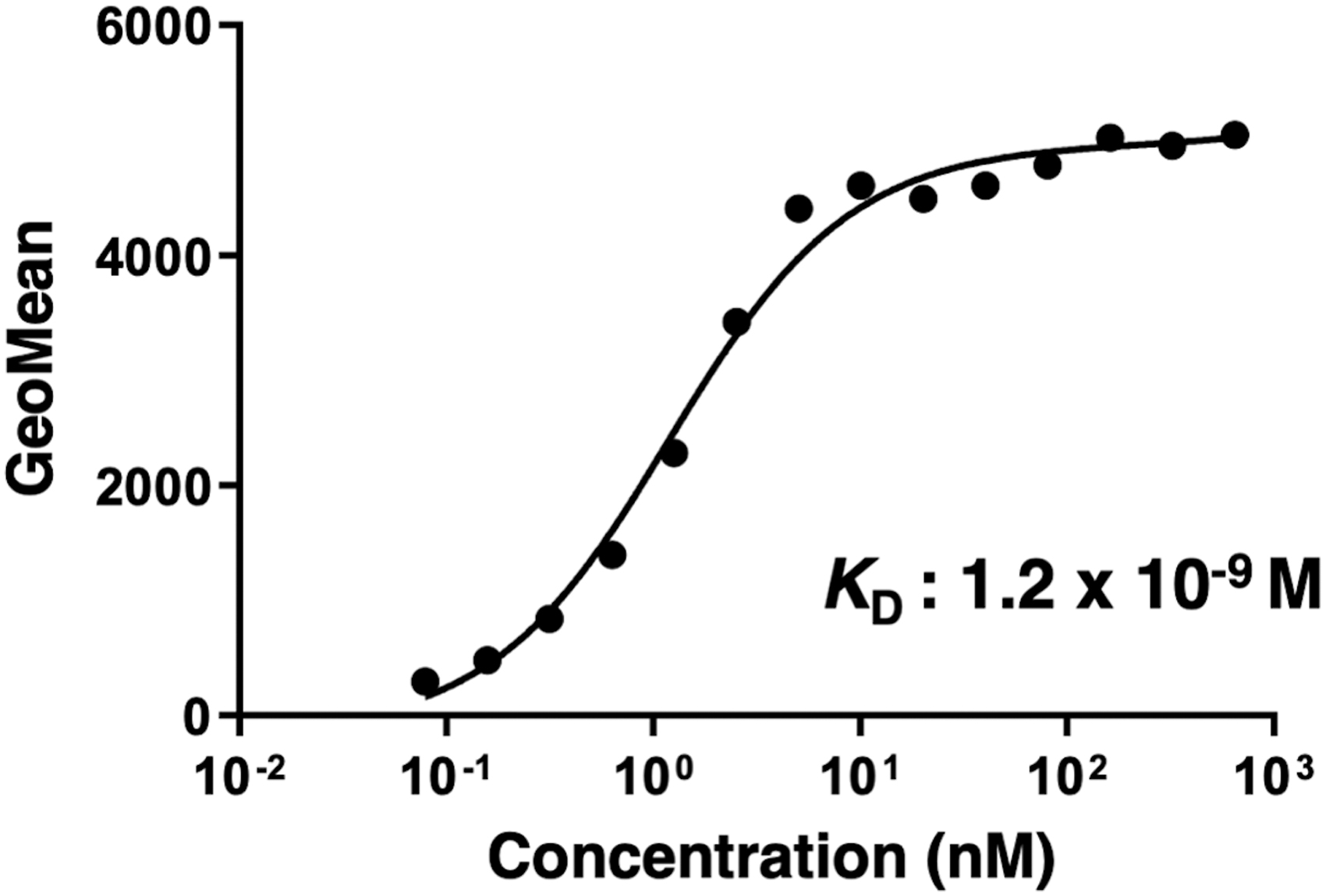
Determination of the binding affinity of C9Mab-1. CHO/hCCR9 was suspended in 100 μL of serially diluted C9Mab-1 (0.006–50 μg/mL). Alexa Fluor 488-conjugated anti-mouse IgG was added. Fluorescence data were collected using the BD FACSLyric.
Discussion
Because the CBIS method does not require purified proteins (Fig. 1), it has been possible to develop antibodies against type I transmembrane and multitransmembrane proteins, such as GPCRs, which are difficult to be purified. We have developed anti-CD20 mAbs(34,35) and anti-CD133 mAbs,(33) and many anti-podoplanin (PDPN) mAbs against pig,(36) Tasmanian devil,(37) alpaca,(38) tiger,(39) whale,(40) goat,(41) horse,(42) and bear(43) PDPNs, anti-PD-L1 mAbs,(44) anti-CD19 mAbs,(45) and anti-CD44 mAbs(46) using the CBIS method. This study established a highly sensitive and specific anti-hCCR9 mAb (clone C9Mab-1) that can be used in flow cytometry using the CBIS method (Fig. 2). Importantly, C9Mab-1 recognized endogenous hCCR9, which is expressed in MOLT-4 cells. Furthermore, the KD of C9Mab-1 for CHO/hCCR9 was 1.2 × 10−9 M (Fig. 3), indicating that C9Mab-1 possesses a high binding affinity for hCCR9.
Chamorro et al.(47) and Somovilla-Crespo et al.(48) previously established an anti-hCCR9 mAb (clone 91R, mouse IgG2b) useful for flow cytometry and Western blotting, and an anti-hCCR9 mAb (clone 92R, mouse IgG2a) useful only for flow cytometry, respectively, by immunizing mice with the eukaryotic expression vector, including full-length hCCR9 using a gene gun. The development of antibodies using this gene gun was first reported by Tang et al.(49) for human growth hormone. This immunization method does not require the adjustment of immunogens and can utilize the post-translational modification system of immunized animals. Therefore, it is possible to develop antibodies that recognize the three-dimensional structure of the target molecule and can be applied to multiple transmembrane proteins.(50–55) However, the immune system might not respond in several conditions: (1) the expression level of the target molecule is low; (2) the target molecule is localized intracellularly; (3) the extracellular region is too small. Also, the gene gun requires expensive equipment.(56)
Several commercially available anti-hCCR9 mAbs were developed using hCCR9-transfected cells as an immunogen without the need for protein purification (Supplementary Table S1). An anti-hCCR9 mAb (clone #112509, mouse IgG2a), used as a positive control in this study, was also immunized using hCCR9-transfected BaF3 mouse proB cell line. Those mAbs are often not applicable to Western blotting because they might recognize the conformation of proteins at the cell membrane, although they are useful for immunohistochemistry or flow cytometry.
In conclusion, we established the anti-hCCR9 mAb, C9Mab-1, applicable to flow cytometry using the CBIS method. The results of our study could also be applied to the more efficient development of other anti-GPCR mAbs.
Footnotes
Authors' Contributions
R.N., J.T., T.A., T.T., M.S., T.N., M.Y., and H.H. performed experiments. M.K.K. designed the experiments. R.N. and Y.K. wrote the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported in part by AMED under Grant Nos. JP20am0401013 (Y.K.) and JP20am0101078 (Y.K.), and by JSPS KAKENHI Grant Nos. 20K16322 (M.S.) and 19K07705 (Y.K.).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.