
Abstract
The development of protein-specific antibodies is essential for understanding a wide variety of biological phenomena. Parasitic and viral infections and cancers are known to occur within California sea lion (Zalophus californianus) populations. However, sensitive and specific monoclonal antibodies (mAbs) for the pathophysiological analysis of California sea lion tissues have not yet been developed. A type I transmembrane glycoprotein, podoplanin (PDPN), is a known diagnostic marker of lymphatic endothelial cells. We have previously developed several anti-PDPN mAbs in various mammalian species, with applications in flow cytometry, Western blotting, and immunohistochemistry. In this study, we established a novel mAb against California sea lion PDPN (seaPDPN), clone PMab-269 (mouse IgG1, kappa), using a Cell-Based Immunization and Screening method. PMab-269 is specifically detected in seaPDPN-overexpressed Chinese hamster ovary (CHO)-K1 cells using flow cytometry and Western blotting. Moreover, PMab-269 clearly identified pulmonary type I alveolar cells, renal podocytes, and colon lymphatic endothelial cells in California sea lion tissues using immunohistochemistry. These findings demonstrate the usefulness of PMab-269 for the pathophysiological analysis of lung, kidney, and lymphatic tissues of the California sea lion.
Introduction
Venous thromboembolism is a life-threatening complication that often occurs in patients with advanced cancer.(1,2) Aberrant activation and aggregation of platelets, and resultant thromboembolisms, have been reported to be associated with cancer progression and metastasis.(3)
Podoplanin (PDPN)/Aggrus/PA2.26 is a type I transmembrane mucin-like sialoglycoprotein involved in platelet aggregation and cancer metastasis through C-type lectin-like receptor-2 (CLEC-2).(4–7) In many cancers, including mesotheliomas,(8,9) oral squamous cell carcinomas (OSCCs),(7,10) and malignant brain tumors,(11–13) PDPN expression is significantly upregulated and the protein is aberrantly glycosylated.(14–16) In normal tissue, PDPN expression is used as a marker of lymphatic endothelial cells, pulmonary type I alveolar cells, and renal podocytes.(17,18) It also plays a pivotal role during embryonic development.(19) Activation of the PDPN-CLEC-2 signaling axis can also trigger infection-induced thrombosis.(20) Higher expression of PDPN significantly reduces the 5-year survival rate of OSCC patients and is linked to advanced-stage tumors.(21) PDPN has also been identified in circulating tumor cells (CTCs), which are defined as a subpopulation of cancer cells harboring high metastatic potential.(22,23) These PDPN-positive CTCs are associated with poorer prognoses in head and neck squamous cell carcinoma patients. Given its potential role in carcinogenesis, PDPN has been exploited as a therapeutic target, with remarkable antitumor effects seen with anti-PDPN monoclonal antibodies (mAbs).(24,25) Recently, anti-PDPN mAbs, such as NZ-1, have been used in chimeric antigen receptor T cell therapy.(26,27) PDPN is also expressed by several cell types within the tumor microenvironment, including inflammatory macrophages, Th17 cells, and cancer-associated fibroblasts (CAFs).(28–30) PDPN-positive CAFs, which are observed in melanoma and non-small-cell lung cancer patients, are correlated with lymph node metastases and poorer prognoses.(31,32)
We have previously developed anti-PDPN mAbs for several mammalian species, including humans,(14,33,34) mice,(35) rats,(36) rabbits,(37) dogs,(38) cats,(39) pigs,(40,41) bovines,(42) Tasmanian devils,(43) alpacas,(44) tigers,(45) goats,(46) horses,(47,48) bears,(49,50) whales,(51) and sheep,(52,53) using Cell-Based Immunization and Screening (CBIS) methods.(54–56) In this study, we successfully developed a novel mAb against California sea lion PDPN (seaPDPN), which can be used for flow cytometry, Western blotting, and immunohistochemistry.
Materials and Methods
Cell lines
Chinese hamster ovary (CHO)-K1 and P3X63Ag8U.1 (P3U1) cells were obtained from the American Type Culture Collection (ATCC, Manassas, VA). Synthesized DNA (Eurofins Genomics KK, Tokyo Japan) encoding seaPDPN (accession no.: XM_027576686) with N-terminal MAP tag(57) or 2 × RIEDL tag(58) was subcloned into a pCAG-Ble vector (FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan). Plasmids were transfected using Lipofectamine LTX with Plus Reagent (Thermo Fisher Scientific, Inc., Waltham, MA). Stable transfectants were selected using cell sorter (SH800; Sony Biotechnology Corp., Tokyo, Japan) and cultivation in media containing 0.5 mg/mL of Zeocin (InvivoGen, San Diego, CA). CHO/MAP-seaPDPN was subsequently used as an immunogen, whereas CHO/2 × RIEDL-seaPDPN was used for further analyses. P3U1, CHO-K1, CHO/MAP-seaPDPN, CHO/2 × RIEDL-seaPDPN, CHO/human PDPN (hPDPN),(5) CHO/mouse PDPN (mPDPN),(5) CHO/rat PDPN (rPDPN),(36) CHO/rabbit PDPN (rabPDPN),(37) CHO/dog PDPN (dPDPN),(38) CHO/bovine PDPN (bovPDPN),(42) CHO/cat PDPN (cPDPN),(39) CHO/pig PDPN (pPDPN),(41) CHO/horse PDPN (horPDPN),(47) CHO/tiger PDPN (tigPDPN),(45) CHO/alpaca PDPN (aPDPN),(44) CHO/bear PDPN (bPDPN),(59) CHO/Tasmanian devil PDPN (tasPDPN),(43) CHO/goat PDPN (gPDPN),(46) CHO/whale PDPN (wPDPN),(51) and CHO/sheep PDPN (sPDPN)(60) were cultured in Roswell Park Memorial Institute (RPMI) 1640 complete media: RPMI 1640 (Nacalai Tesque, Inc., Kyoto, Japan) supplemented with 10% heat-inactivated fetal bovine serum (Thermo Fisher Scientific, Inc.), 100 U/mL of penicillin, 100 μg/mL streptomycin, and 0.25 μg/mL amphotericin B (Nacalai Tesque, Inc.), and incubated at 37°C in a humidified atmosphere containing 5% CO2.
Antibodies
Mouse anti-hPDPN mAb LpMab-12 (IgG1, kappa),(61) rat anti-mPDPN mAb PMab-1 (IgG2a, kappa),(35) mouse anti-rPDPN mAb PMab-2 (IgG1, kappa),(36) mouse anti-rabPDPN mAb PMab-32 (IgG1, kappa),(37) mouse anti-dPDPN mAb PMab-38 (IgG1, kappa),(38) mouse anti-bovPDPN mAb PMab-44 (IgG1, kappa),(42) mouse anti-cPDPN mAb PMab-52 (IgM, kappa),(39) mouse anti-pPDPN mAb PMab-213 (IgG2b, kappa),(40) mouse anti-horPDPN mAb PMab-219 (IgG2a, kappa),(62) mouse anti-tasPDPN mAb PMab-233 (IgG1, kappa),(43) mouse anti-gPDPN mAb PMab-235 (IgG1, kappa),(46) mouse anti-aPDPN mAb PMab-225 (IgG2b, kappa),(44) mouse anti-tigPDPN mAb PMab-231 (IgG2a, kappa),(45) mouse anti-bPDPN mAb PMab-247 (IgG1, kappa),(59) mouse anti-wPDPN mAb PMab-237 (IgG1, kappa),(51) and mouse anti-sPDPN mAb PMab-256 (IgG1, kappa)(52) were developed as previously described.
Hybridoma production
Female BALB/c mice (6-week-old) were purchased from CLEA Japan (Tokyo, Japan). The animals were housed under specific pathogen-free conditions. The Animal Care and Use Committee of Tohoku University approved all animal experiments. We used a CBIS method to develop mAbs against seaPDPN. A BALB/c mouse was immunized with CHO/MAP-seaPDPN cells (1 × 108) intraperitoneally with Imject Alum (Thermo Fisher Scientific, Inc.). The procedure included three additional immunizations with CHO/MAP-seaPDPN cells (1 × 108) followed by a final booster injection of CHO/MAP-seaPDPN cells (1 × 108) 2 days before the harvest of splenic cells. Subsequently, splenic cells were fused with P3U1 cells using polyethylene glycol 1500 (PEG1500; Roche Diagnostics, Indianapolis, IN). The hybridomas were then grown in RPMI complete media supplemented with hypoxanthine, aminopterin, and thymidine for selection (Thermo Fisher Scientific, Inc.). The culture supernatants were screened for anti-seaPDPN antibodies production using flow cytometry.
Flow cytometry
CHO/2 × RIEDL-seaPDPN and parental CHO-K1 cells (2 × 105/mL) were harvested after a brief exposure to 0.25% trypsin in 1 mM ethylenediaminetetraacetic acid (EDTA; Nacalai Tesque, Inc.). After washing with 0.1% bovine serum albumin (Nacalai Tesque, Inc.) in phosphate-buffered saline (PBS; Nacalai Tesque, Inc.), cells were treated with anti-PDPN mAbs (5 μg/mL) or an anti-RIEDL tag (clone LpMab-7; 10 μg/mL) for 30 minutes at 4°C, followed by Alexa Fluor 488-conjugated anti-mouse IgG (1:1000, product no. 4408; Cell Signaling Technology, Inc., Danvers, MA) or Alexa Fluor 488-conjugated anti-rat IgG (1:1000; product no. 4416; Cell Signaling Technology, Inc.). Fluorescence data were collected using the SA3800 Cell Analyzer (Sony Biotechnology Corp.).
Western blot analysis
Cell lysates (10 μg) were boiled in sodium dodecyl sulfate (SDS) sample buffer (Nacalai Tesque, Inc.). Proteins were electrophoresed on 5%–20% polyacrylamide gels (FUJIFILM Wako Pure Chemical Corporation) and transferred onto polyvinylidene difluoride (PVDF) membranes (Merck KGaA, Darmstadt, Germany). After blocking with 4% skim milk (Nacalai Tesque, Inc.), PVDF membranes were incubated with 5 μg/mL of PMab-269, 1 μg/mL of anti-RIEDL tag (clone LpMab-7), or 1 μg/mL of anti-β-actin (clone AC-15; Sigma-Aldrich Corp., St. Louis, MO), followed by incubation with peroxidase-conjugated anti-mouse immunoglobulins (diluted 1:1000; Agilent Technologies, Inc., Santa Clara, CA). Blots were developed using ImmunoStar LD (FUJIFILM Wako Pure Chemical Corporation) and imaged with a Sayaca-Imager (DRC Co. Ltd., Tokyo, Japan).
Immunohistochemical analysis
Normal California sea lion tissues obtained from routine necropsies performed at the Laboratory of Veterinary Pathology, the University of Tokyo, fixed in 10% neutral-buffered formalin, were processed to make formalin-fixed paraffin-embedded (FFPE) tissue sections.(63) Histological sections (4 μm thickness) were autoclaved in citrate buffer (pH 6.0; Nichirei Biosciences, Inc., Tokyo, Japan) or EnVision FLEX Target Retrieval Solution High pH (Agilent Technologies, Inc.) for 20 minutes. After blocking with SuperBlock T20 (PBS) Blocking Buffer (Thermo Fisher Scientific, Inc.), sections were incubated with PMab-269 (5 μg/mL) for 1 hour at room temperature and treated using EnVision+ Kit (Agilent Technologies, Inc.) for 30 minutes. Color was developed using 3,3′-diaminobenzidine tetrahydrochloride (DAB; Agilent Technologies, Inc.) for 2 minutes, and counterstaining was performed with hematoxylin and eosin (FUJIFILM Wako Pure Chemical Corporation).
Results
Establishment of anti-seaPDPN mAbs
To develop anti-seaPDPN mAbs, we used the CBIS method, using stable transfectants for immunization and flow cytometry (Fig. 1). A mouse was immunized with CHO/MAP-seaPDPN cells, which overexpress seaPDPN with an N-terminal MAP tag. Hybridomas were seeded into 96-well plates, and CHO/2 × RIEDL-seaPDPN (hereinafter referred to as CHO/seaPDPN)-positive and CHO-K1-negative wells were identified. After limiting dilution and selection using Western blotting and immunohistochemistry, PMab-269 (mouse IgG1, kappa) was isolated.
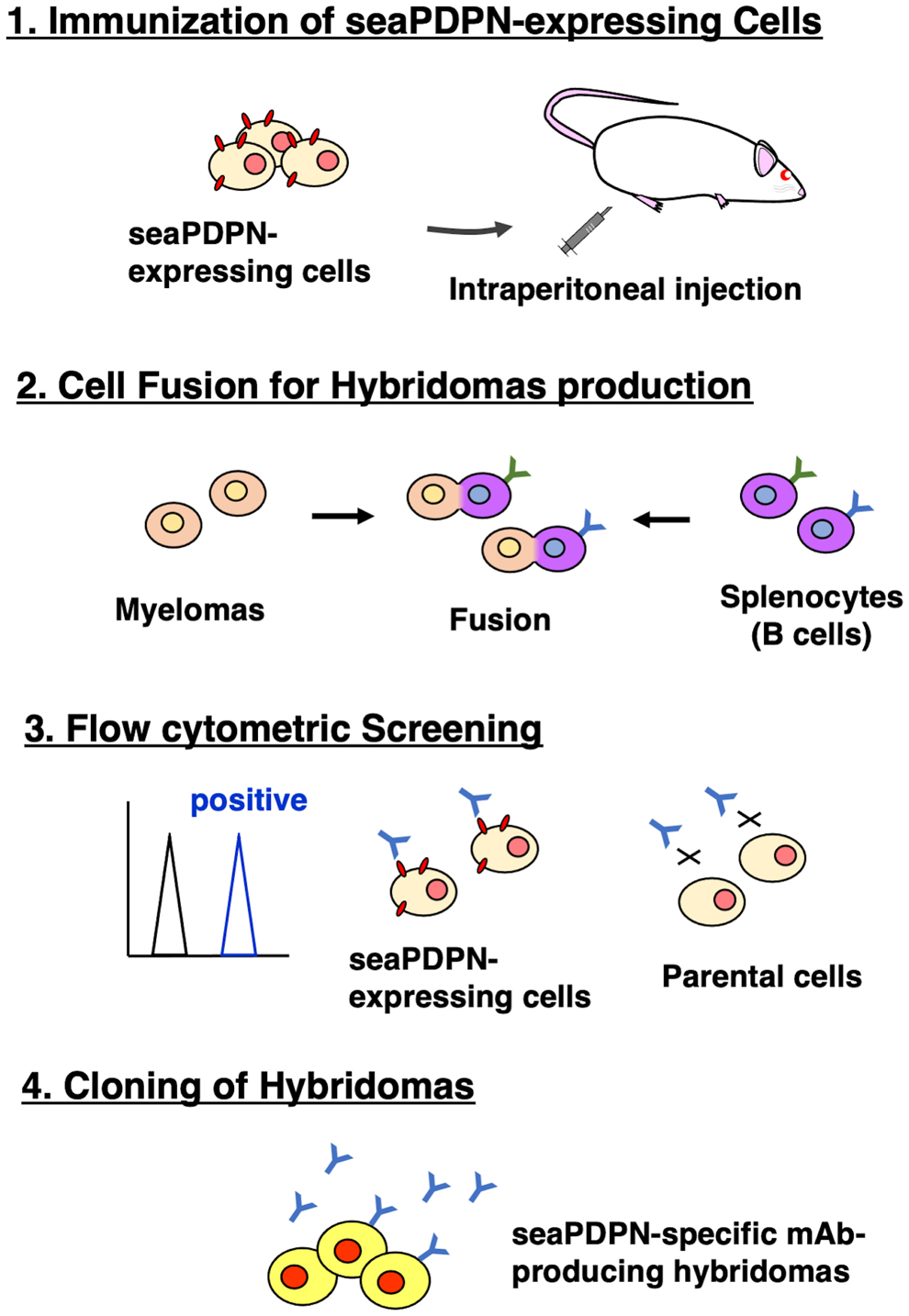
Schematic model of the CBIS method. seaPDPN-overexpressed stable transfectants were used as an immunogen. Anti-seaPDPN mAb-secreting hybridomas were screened for using flow cytometry of parental cells and stable transfectants. CBIS, Cell-Based Immunization and Screening; mAb, monoclonal antibody; seaPDPN, sea lion podoplanin.
Flow cytometric analyses
We performed flow cytometric analyses using PMab-269 with CHO/seaPDPN and CHO-K1 cells. PMab-269 recognized CHO/seaPDPN but not CHO-K1 cells (Fig. 2A). As expected, an anti-RIEDL tag mAb (LpMab-7) could also detect CHO/seaPDPN cells (Fig. 2B). These results demonstrate that PMab-269 is suitable the detection of seaPDPN using flow cytometry.
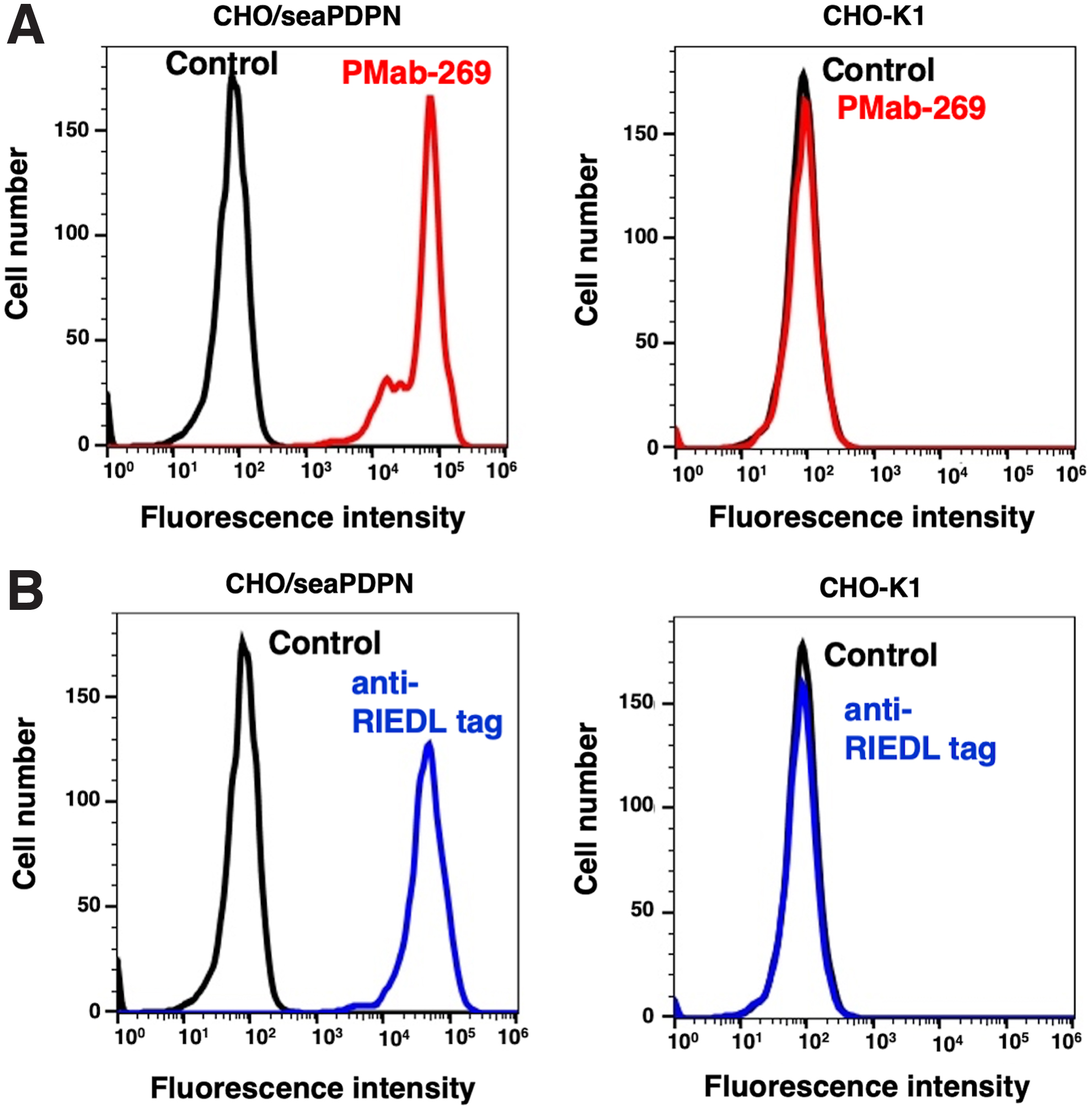
Detection of seaPDPN using PMab-269 through flow cytometry.
We evaluated crossreactivity of PMab-269 with other animal PDPNs, which are overexpressed in CHO-K1 cells. PMab-269 crossreacted with rat, dog, cat, alpaca, and tiger PDPNs (Fig. 3A). The expression levels of each of these PDPNs were confirmed using their specific anti-PDPN mAbs (Fig. 3B).
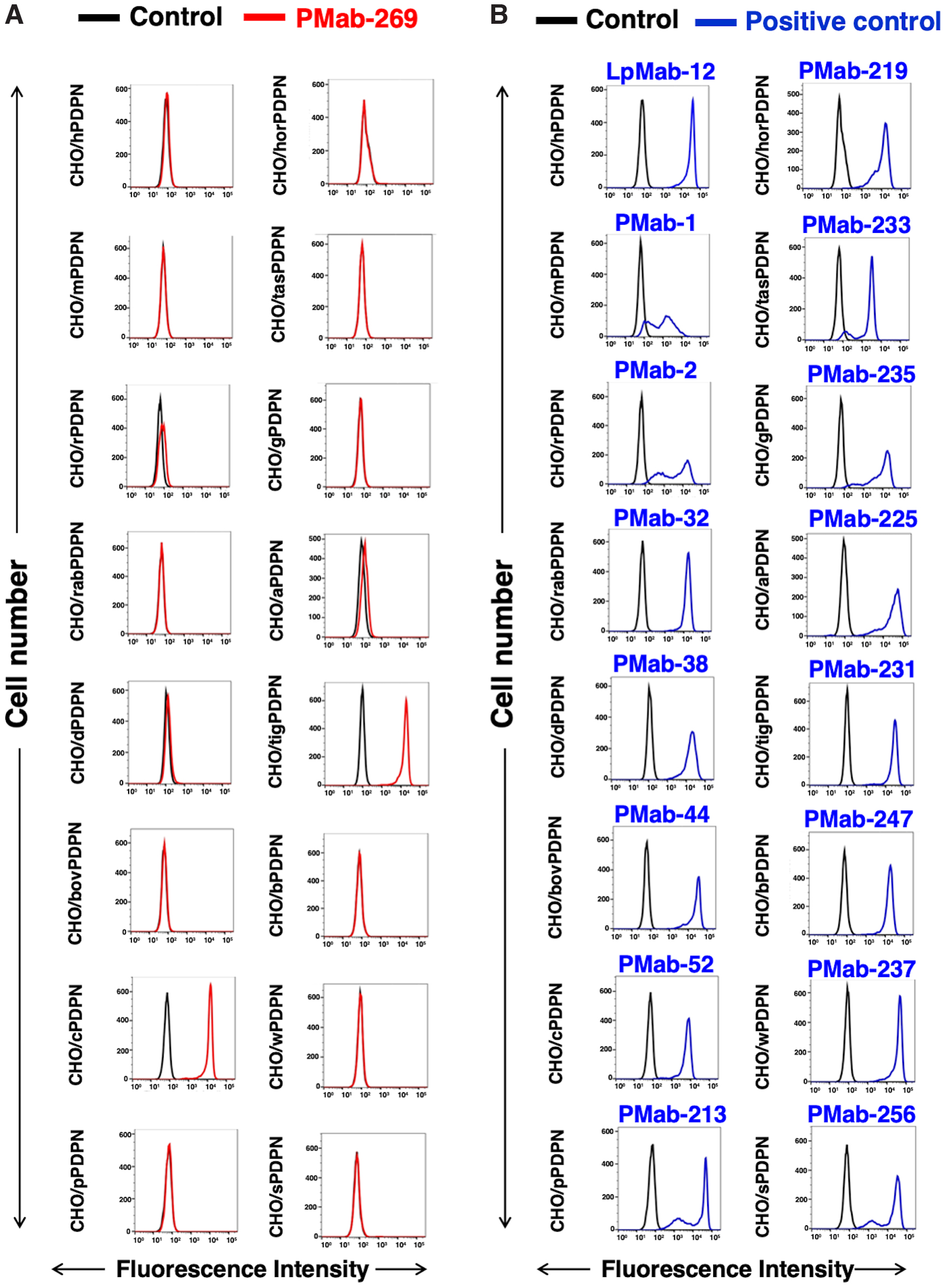
Crossreactivity of PMab-269 with PDPNs of other species.
Western blot analyses
We investigated whether PMab-269 can be used for Western blot analyses by analyzing CHO-K1 and CHO/seaPDPN cell lysates. As shown in Figure 4, PMab-269 could detect seaPDPN as a 40 kDa band (2 × RIEDL+seaPDPN) in CHO/seaPDPN cell lysates, whereas no major bands were detected in the parental CHO-K1 cells. Anti-RIEDL tag (LpMab-7) was used as a positive control and could also detect a band of the same molecular weight in CHO/seaPDPN cell lysates. The 40 kDa major band represents a full-length form, whereas the lower band (35 kDa), which was detected by both PMab-269 and LpMab-7, is presumably a truncated isoform.(5,14) Additionally, two weaker, smaller bands (35–28 kDa) and two larger bands (100–63 kDa) were detected, which may represent additional truncated and aggregated isoforms, respectively. These results indicate that PMab-269 can detect seaPDPN in Western blot analyses.
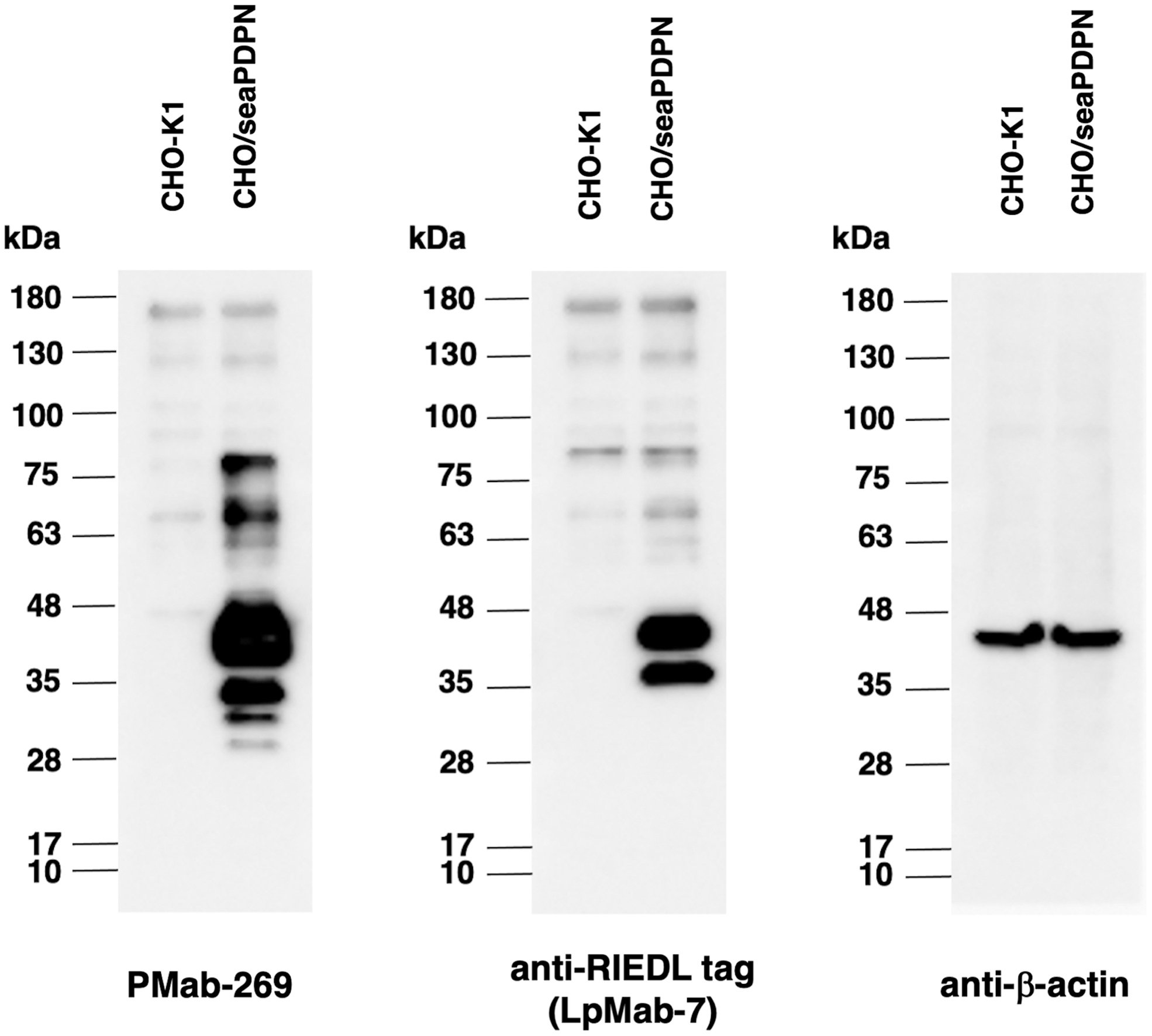
Detection of seaPDPN by Western blot analysis. Cell lysates of CHO-K1 and CHO/seaPDPN (10 μg) were electrophoresed and transferred onto PVDF membranes. The membranes were incubated with 5 μg/mL of PMab-269, 1 μg/mL of an anti-RIEDL tag mAb (LpMab-7), and an anti-β-actin mAb (AC-15) and subsequently with peroxidase-conjugated anti-mouse immunoglobulins. PVDF, polyvinylidene difluoride.
Immunohistochemical analyses
Lastly, we investigated whether PMab-269 can be used for immunohistochemical analyses using FFPE California sea lion tissue sections. We examined normal lung, kidney, and colon tissue from California sea lions, all of which reportedly express PDPN in various species, including humans,(33) mice,(35) rats,(36) rabbits,(37) dogs,(38) cats,(39) bovines,(42) pigs,(40) Tasmanian devils,(43) alpacas,(44) tigers,(45) whales,(51) goats,(46) horses,(47) bears,(49) and sheep.(52,53) As shown in Figure 5, PMab-269 strongly and specifically identified type I alveolar cells within the lung. PMab-269 also identified renal glomeruli and Bowman's capsules (Fig. 6). Furthermore, lymphatic endothelial cells of colon were also detected by PMab-269 (Fig. 7). These results indicate that PMab-269 is useful in the detection of seaPDPN-positive cells in FFPE tissue.
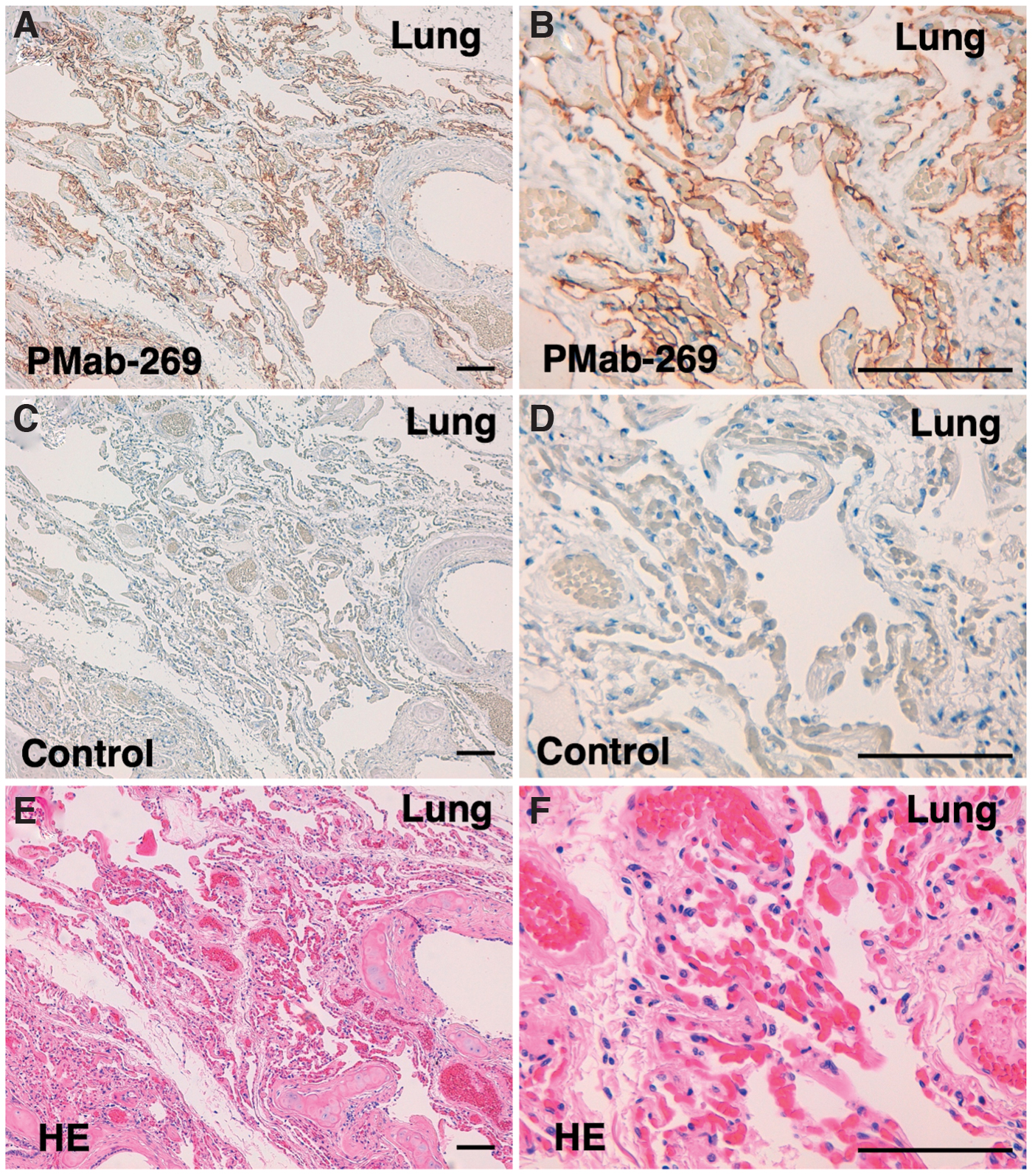
Immunohistochemical analysis of California sea lion lung tissue. Histological sections of California sea lion lung were directly autoclaved in citrate buffer and incubated with 5 μg/mL of PMab-269
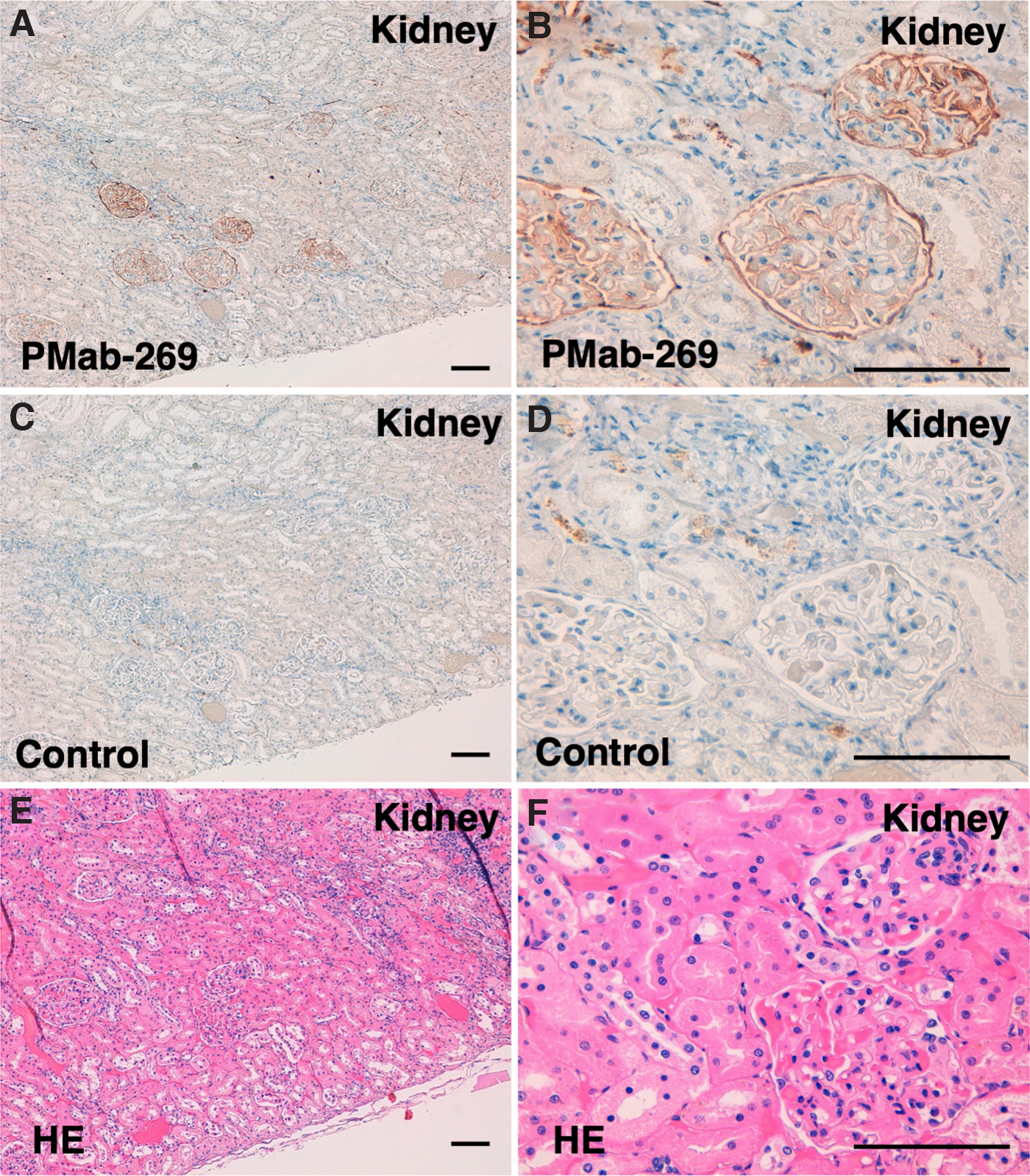
Immunohistochemical analysis of California sea lion kidney tissue. Histological sections of California sea lion kidney were directly autoclaved in EnVision FLEX Target Retrieval Solution High pH and incubated with 5 μg/mL of PMab-269
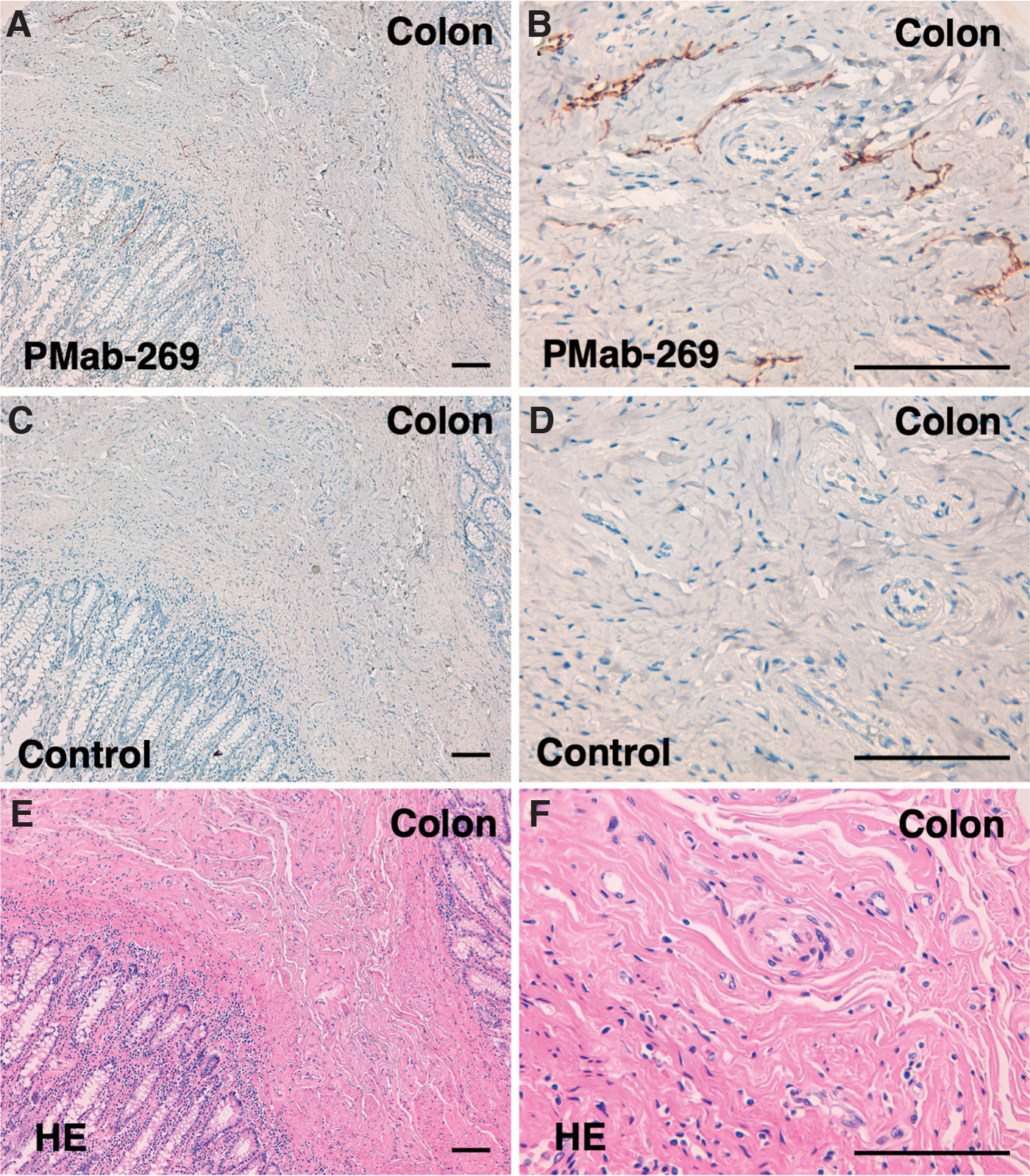
Immunohistochemical analyses of California sea lion colon tissue. Histological sections of California sea lion colon were directly autoclaved in citrate buffer and incubated with 5 μg/mL of PMab-269
Discussion
Animal models of pathological conditions are essential for elucidating pathogenesis and developing effective therapeutic agents. Although California sea lion (Zalophus californianus) is a nonlaboratory animal, infectious diseases, such as parasites and viruses, and several cancers, including metastatic carcinoma of urogenital origin, intestinal T cell lymphoma, and lingual squamous cell carcinoma, have been observed among their populations.(64–67) In addition to the carcinogenesis observed in women infected with the papillomavirus, a relationship between carcinogenesis and herpesvirus and bacterial infections has similarly been observed in California sea lion.(68) Immune system analyses and genetic research of California sea lions could thus improve our understanding of human pathology.(66,69)
Although pathological analyses, such as immunohistochemistry, have been previously reported, specific antibodies against California sea lion proteins are rarely used.(63,70) In previous studies, the expression of California sea lion proteins, including the estrogen receptor, progesterone receptor, amyloid-β and tau, have been evaluated through immunohistochemistry by exploiting antibodies raised in other species. However, such approaches may limit sensitivity and specificity. In this study, we developed a novel mAb (PMab-269) against seaPDPN using the CBIS method (Fig. 1). PMab-269 could detect seaPDPN by flow cytometry (Fig. 2), Western blotting (Fig. 4), and immunohistochemistry (Figs. 5–7). In future studies, PMab-269 could be used to obtain pathophysiological data for California sea lions.
Footnotes
Authors' Contributions
T.T., T.A., M.S., J.T., H.Ho, R.N, T.N., M.Y., and M.Y. performed experiments; M.K.K., H.Ha, M.F., H.S., K.U., and T.N. designed the experiments; T.T. and Y.K. wrote the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported in part by Japan Agency for Medical Research and Development (AMED) under Grant Numbers: JP21am0401013 (to Y.K.) and JP21am0101078 (to Y.K.), and by the Japan Society for the Promotion of Science (JSPS) Grants-in-Aid for Scientific Research (KAKENHI) grant nos. 21K15523 (to T.A.), 21K07168 (to M.K.K.), 20K16322 (to M.S.), and 19K07705 (to Y.K.).