
Abstract
CD44 functions as a major hyaluronan receptor on most cell types, with roles in cell adhesion, migration, proliferation, differentiation, and survival. The CD44 gene comprises 20 exons, with alternative splicing producing many different isoforms. CD44 variant isoforms exhibit tissue-specific expression patterns and have been studied as therapeutic targets for several cancers; therefore, anti-CD44 monoclonal antibodies (mAbs) are useful for investigating CD44 expression in various cancers. Previously, we established an anti-CD44 mAb, C44Mab-46 (IgG1, κ), by immunizing mice with the CD44v3–10 ectodomain. Although C44Mab-46 recognized all CD44 isoforms, the binding epitope of C44Mab-46 has not been determined. In this study, we first checked the reactivity of C44Mab-46 to several CD44v3–10 deletion mutants such as dN79, dN124, dN147, and dN224. We found the N-terminus of the C44Mab-46-binding epitope between residues 147 and 224 of CD44v3–10. We next investigated this epitope using a novel mapping system: RIEDL insertion for epitope mapping (REMAP) method. We constructed 31 CD44 standard (CD44s) mutants where the RIEDL tag was inserted into the expected epitope region in CD44s. We observed that the C44Mab-46 epitope constituted five amino acids: 174-TDDDV-178 of CD44s. Thus, the REMAP method could be used to determine mAb binding epitopes for membrane proteins.
Introduction
CD44 is a type I transmembrane glycoprotein widely expressed on the surface of many cell types, including epithelial, fibroblast, and leukocyte cells.(1) CD44 functions as a hyaluronan (HA) receptor, with CD44–HA interactions mediating cell adhesion and migration in several physiological and pathological processes.(2,3) The CD44 structure comprises extracellular, transmembrane, and intracellular domains. The extracellular domain contains an HA-binding region(4) and an alternative splicing site. The CD44 gene comprises 20 exons, with alternative splicing producing many different isoforms, such as CD44 standard (CD44s) that lacks variant exons, and CD44v3–10, with exons v3 to v10.(5) The intracellular domain contains two ligand binding sites: an ezrin, radixin, and moesin (ERM) binding site and an ankyrin binding site. CD44 links HA and the membrane cytoskeleton via these binding sites to ERM and ankyrin.(6–9) Moreover, the intracellular domain of CD44 is also necessary for the binding of soluble HA; therefore, this domain potentially mediates ligand binding at the extracellular domain.(10)
CD44 is related to development, metastasis, and tumor invasion(11–14); therefore, it is a biomarker and therapeutic target in several cancers.(15) We previously developed several anti-CD44 monoclonal antibodies (mAbs),(16) including clone C44Mab-46, by immunizing mice with the CD44v3–10 ectodomain. C44Mab-46 recognized not only CD44s but also CD44 variants. However, the C44Mab-46 epitope remains undetermined.
To determine molecular epitopes, several methods may be used.(17) X-ray cocrystallography allows the direct visualization of interactions between antigens and antibodies; however, the approach takes time and is expensive. By contrast, array-based oligopeptide scanning and site-directed mutagenesis mapping can easily determine linear epitopes, but these methods are inappropriate for conformational epitopes. Recently, we developed a novel epitope mapping system: the RIEDL insertion for epitope mapping (REMAP) method using the RIEDL tag system.(18) This approach is simple and efficient for the determination of conformational epitopes. In this study, we identified the epitope of C44Mab-46 for CD44 using the REMAP method.
Materials and Methods
Plasmid construction
The CD44s open reading frame (ORF) was amplified from LN229 complementary DNA using the HotStar HiFidelity Polymerase Kit (Qiagen Inc., Hilden, Germany).(16) The CD44v3–10 ORF was provided by the RIKEN BioResource Research Center via the National Bioresource Project of MEXT, Japan.(16) Both CD44s and CD44v3–10 ORFs were separately subcloned into the pCAG-Ble vector (FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan), and a signal sequence (MYRMQLLSCIALSLALVTNS) and PA16 tag (GLEGGVAMPGAEDDVV),(16,19–23) which is recognized by an anti-PA16 tag mAb (NZ-1),(24) were added at each N-terminus. CD44v3–10 deletion mutants were constructed using the HotStar HiFidelity Polymerase Kit (Qiagen Inc.), using oligonucleotides containing the desired mutations. Insertion of RIEDL tags into CD44s sequences was performed with oligonucleotides containing RIEDL tag insertions at the desired position. For example, by inserting the RIEDL sequence (R*) between Glu160 and Tyr161 of CD44s, we constructed Glu160_RIEDL_Tyr161 (E160_R*_Y161). PCR fragments bearing the desired mutations were inserted into pCAG-Ble using an In-Fusion HD Cloning Kit (Takara Bio, Inc., Shiga, Japan). The RIEDL tag insertion mutants were as follows: E160_R*_Y161, Y161_R*_R162, R162_R*_T163, T163_R*_N164, N164_R*_P165, P165_R*_E166, E166_R*_D167, D167_R*_I168, I168_R*_Y169, Y169_R*_P170, P170_R*_S171, S171_R*_N172, N172_R*_P173, P173_R*_T174, T174_R*_D175, D175_R*_D176, D176_R*_D177, D177_R*_V178, V178_R*_S179, S179_R*_S180, S180_R*_G181, G181_R*_S182, S182_R*_S183, S183_R*_S184, S184_R*_E185, E185_R*_R186, R186_R*_S187, S187_R*_S188, S188_R*_T189, T189_R*_S190, and S190_R*_G191.
Cell lines
Chinese hamster ovary (CHO)-K1 cells were purchased from the America Type Culture Collection (ATCC, Manassas, VA). CD44 mutant plasmids were transfected into cells using the Neon Transfection System (Thermo Fisher Scientific, Inc., Waltham, MA), and stable transfectants were sorted using NZ-1 with a cell sorter (SH800; Sony Corp., Tokyo, Japan). Cells were cultured in the RPMI 1640 medium (Nacalai Tesque, Inc., Kyoto, Japan), supplemented with 10% heat-inactivated fetal bovine serum (Thermo Fisher Scientific Inc.), 100 U/mL penicillin, 100 μg/mL streptomycin, and 0.25 μg/mL amphotericin B (Nacalai Tesque, Inc.) at 37°C in a humidified atmosphere containing 5% CO2. Transfectants were cultivated in a medium containing 0.5 mg/mL Zeocin (InvivoGen, San Diego, CA).
Flow cytometry
Cells were harvested by brief exposure to 0.25% trypsin/1 mM ethylenediaminetetraacetic acid (Nacalai Tesque, Inc.). After washing in 0.1% bovine serum albumin in phosphate-buffered saline (blocking buffer), cells were treated with primary mAbs, such as C44Mab-46 (mouse IgG1, kappa), NZ-1 (rat IgG2a, lambda), or LpMab-7 (mouse IgG1, kappa) at a concentration of 1 μg/ml for 30 minutes at 4℃; subsequently, with Alexa Fluor 488-conjugated anti-mouse IgG or Alexa Fluor 488-conjugated anti-rat IgG (1:1000; Cell Signaling Technology, Inc., Danvers, MA). Fluorescence data were collected using a BD FACSLyric (Becton, Dickinson and Company, Franklin Lakes, NJ) instrument for deletion mutant analysis, or the EC800 Cell Analyzer (Sony Corp.) for REMAP analyses.
Results
Determination of the C44Mab-46 epitope using CD44v3–10 deletion mutants
The CD44 gene comprises 20 exons. The smallest isoform is the standard form, CD44s, and other variant isoforms include CD44v3–10 (Fig. 1A). The CD44 extracellular domain is encoded by exons 1–17. First, we constructed CD44v3–10 N-terminal deletion mutants (dN79, dN124, dN147, and dN224), with the PA16 tag at their N-terminus. Reactions between C44Mab-46 and deletion mutants were characterized by flow cytometry and showed that C44Mab-46 recognized CD44v3–10, dN79, dN124, and dN147, but not dN224 (Fig. 1B). By contrast, all deletion mutants were detected by the anti-PA16 tag mAb, NZ-1 (Fig. 1C). C44Mab-46 and NZ-1 did not recognize CHO-K1 parental cells (Figs. 1B, C). These results indicated that the N-terminus of the C44Mab-46 epitope existed between the 147th and 224th amino acids of CD44v3–10 (Fig. 1D).
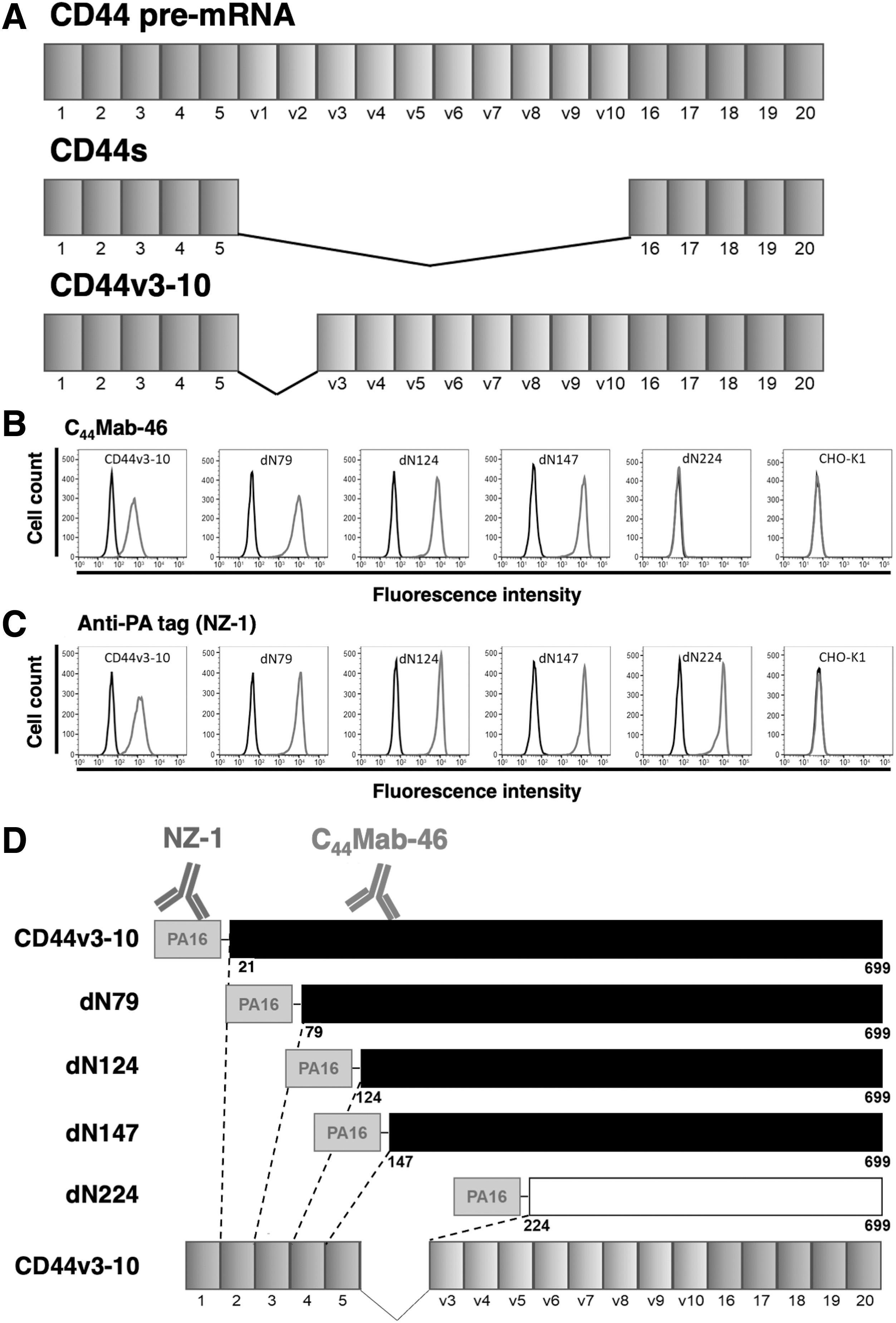
Epitope mapping of C44Mab-46 using CD44v3–10 deletion mutants.
Determination of the C44Mab-46 epitope using REMAP method
To further investigate the C44Mab-46 epitope, we performed the REMAP method. For this, we used the RIEDL tag system,(18) which comprised a five-amino acid peptide (RIEDL tag) and an anti-RIEDL tag mAb (clone LpMab-7). C44Mab-46 bound to CD44s when the RIEDL tag was inserted into any region, which was independent of the C44Mab-46 epitope (Fig. 2A, upper panel). By contrast, CD44s was not detected by C44Mab-46 when C44Mab-46 epitope conformation was disrupted by RIEDL insertion (Fig. 2A, lower panel). We constructed 31 CD44s mutants where the RIEDL tag was inserted into the expected epitope region in CD44s (Fig. 2B). Our flow cytometry results showed that C44Mab-46 did not detect four mutants (T174_R*_D175, D175_R*_D176, D176_R*_D177, and D177_R*_V178) and weakly detected P173_R*_T174 (Fig. 3A), suggesting C44Mab-46 bound to CD44s at five amino acids (from Thr174 to Val178). As positive controls, NZ-1 against the N-terminal PA16 tag and LpMab-7 against the RIEDL tag detected all mutants (Fig. 3B, C). Since the RIEDL tag was not inserted into wild-type CD44s, LpMab-7 did not react with CHO/CD44s (Fig. 3C). These results are summarized (Fig. 4).

RIEDL insertion for the REMAP method. RIEDL tags were inserted into CD44s.
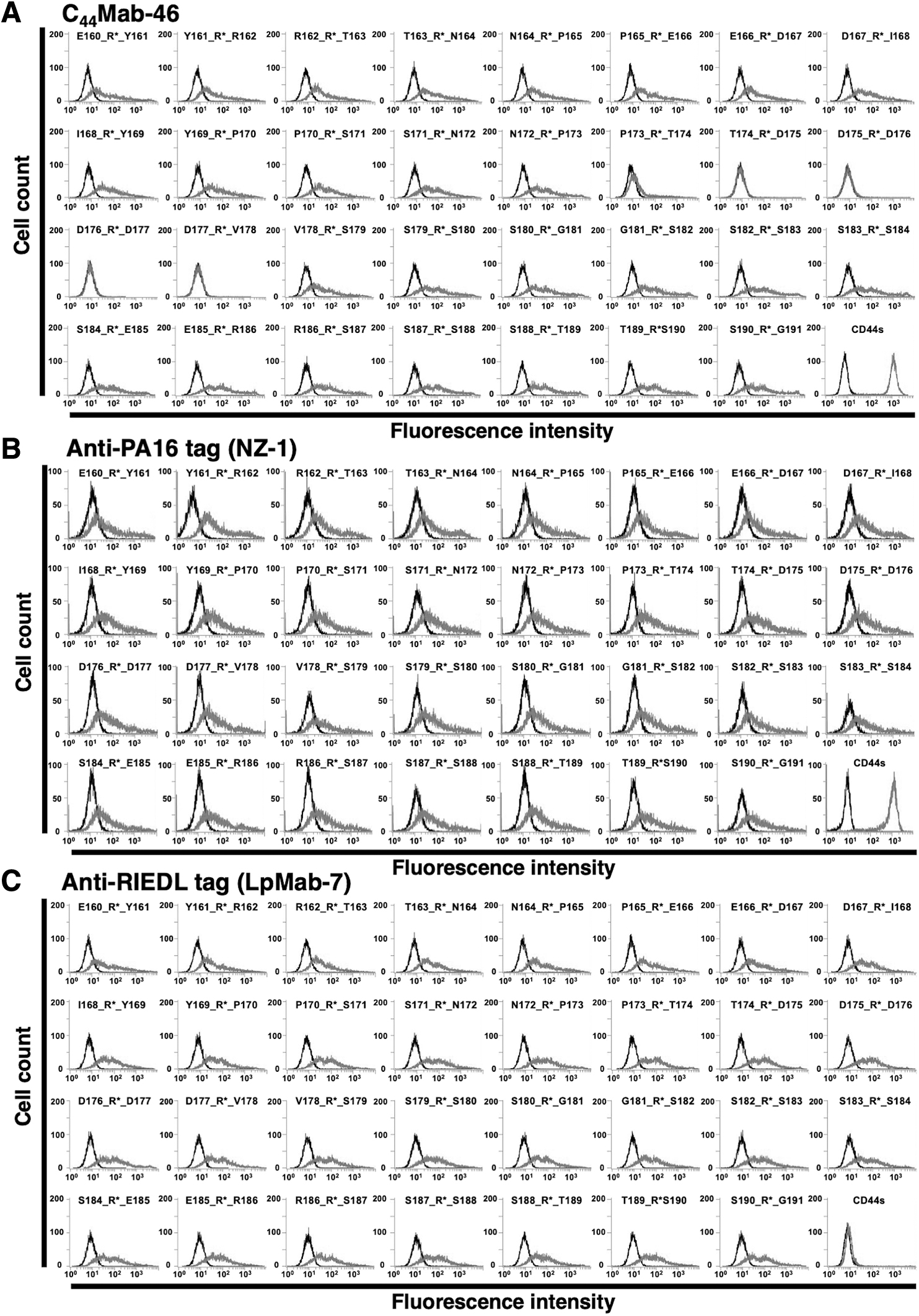
Epitope mapping of C44Mab-46 using RIEDL-tagged CD44s insertion mutants. RIEDL tags were inserted into CD44s, with mutants analyzed by flow cytometry. RIEDL tag insertion mutants were expressed in CHO-K1 cells and incubated with
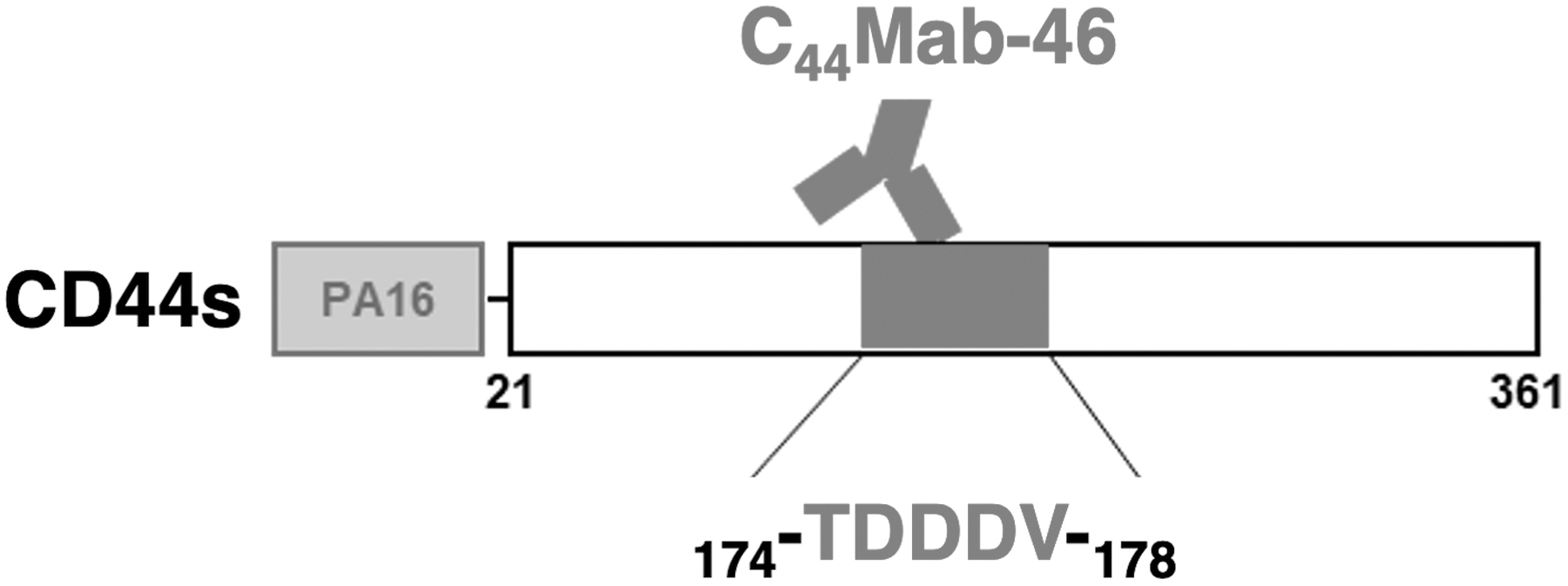
Summary of epitope mapping of C44Mab-46. Thr174, Asp175, Asp176, Asp177, and Val178 are critical amino acids for C44Mab-46 binding to CD44s.
Discussion
To investigate mAb binding epitopes, alanine-scanning mutagenesis and peptide screening methodologies are generally used.(17) These methods are simple and effective for identifying linear epitopes; however, they are unsuited to conformational epitopes. To determine these epitopes, we developed the REMAP method.(18)
We previously developed several anti-CD44 mAbs, including C44Mab-46 and C44Mab-5 by immunizing mice with CD44v3–10 ectodomain and CD44v3-10-overexpressed CHO-K1 cells, respectively.(16) To investigate the C44Mab-46 epitope, we conducted N-terminal deletion mutant analyses. Our data showed that the N-terminus of the C44Mab-46 epitope was located between amino acids 147 and 224 of CD44v3–10 (Fig. 1). This epitope was also located in CD44s. Using additional deletion mutants, we further found that the C44Mab-46 epitope was located between amino acids 161 and 190 of CD44s (data not shown). To identify critical amino acids, we further used the REMAP method (Fig. 2). Flow cytometry analyses showed that NZ-1 and LpMab-7 recognized all RIEDL tag insertion mutants. By contrast, C44Mab-46 lost its reactivity to four mutants of CD44s (Fig. 3). RIEDL tag insertion may disrupt the structure of the epitope region and inhibit the binding of C44Mab-46 to CD44s. We successfully determined the C44Mab-46-binding epitope for CD44s as 174-TDDDV-178 (Fig. 4). These amino acids were encoded in exon 5 (Fig. 1); therefore, C44Mab-46 could detect not only CD44s but also all CD44 variant isoforms. Identifying the C44Mab-46 epitope will help in the development of a CD44-targeting therapeutic antibody in the future.
LpMab-7 can detect the RIEDL tag, which is inserted into the loop structure of protein.(18) Therefore, the REMAP method could be advantageous for determination of the conformational epitope of various membrane proteins, even if the epitope cannot not be determined by the alanine scanning method in the future study.
Footnotes
Authors' Contributions
T.A., J.T., and N.T. performed experiments; M.K.K. designed the experiments; and T.A. and Y.K. wrote the article.
Author Disclosure Statement
The authors have no conflict of interest.
Funding Information
This research was supported, in part, by the Japan Agency for Medical Research and Development (AMED) under grant nos: JP21am0401013 (Y.K.) and JP21am0101078 (Y.K.), and by the Japan Society for the Promotion of Science (JSPS) Grants-in-Aid for Scientific Research (KAKENHI) grant nos. 21K15523 (to T.A.), 21K07168 (to M.K.K.), and 19K07705 (to Y.K.).