
Abstract
Podoplanin (PDPN) plays a pivotal role in platelet aggregation, embryo development, and tumor progression. PDPN is universally expressed in many mammalian species, and is considered a typical lymphatic endothelial cell marker. We have previously developed the mouse anti-California sea lion (Zalophus californianus) PDPN (seaPDPN) monoclonal antibody (mAb), clone PMab-269, which is suitable for different experimental applications, including flow cytometry, Western blotting, and immunohistochemistry. In this study, we identified the PMab-269 epitope of the seaPDPN by enzyme-linked immunosorbent assay using deletion mutants and point mutants generated for seaPDPN. Our results demonstrated that PMab-269 recognized the peptide, corresponding to the amino acids 63–82 of seaPDPN. Furthermore, the reactions of PMab-269 to seven alanine-substituted peptides, such as P68A, D76A, F77A, H78A, L79A, E80A, and D81A, were abolished among 20 alanine-substituted peptides. We identified the seven amino acids (Pro68, Asp76, Phe77, His78, Leu79, Glu80, and Asp81) as the critical epitope targeted by PMab-269. The successful identification of the PMab-269 epitope might contribute to the pathophysiological investigations of seaPDPN.
Introduction
Epitope identification and design of immunogen are essential for the development of monoclonal antibodies (mAbs) and vaccines.(1,2) It is important to determine whether an epitope recognized by a mAb can be used for the development of tag systems for protein extraction, purification, or detection assays.(3–8) Application of epitope tag systems facilitates the detection of membrane proteins that are structurally difficult to recognize by antibodies alone.
Podoplanin (PDPN), also known as Aggrus or PA2.26, is a single pass transmembrane glycoprotein that is highly expressed in normal tissues, such as lymphatic endothelial cells, type I alveolar cells, and renal glomeruli in mammals.(9–11) The amino acids sequence of PDPN is well conserved across mammals.(12) PDPN is involved in platelet aggregation by binding to its receptor, the C-type lectin-like receptor-2 (CLEC-2).(13) The platelet aggregation–stimulating (PLAG) domain, EDxxVTPG sequence, and the PLAG-like domain (PLD), E(D/E)xx(T/S)xx sequence, are critical for the platelet-aggregating activity of PDPN.(12,14–16) PDPN–CLEC-2 interaction is also important for blood/lymphatic vascular separation in the embryo.(17) In the immune system, PDPN is known to influence the maturation of natural regulatory T cells and antigen-presenting dendritic cell migration to lymphatic vessels through interactions with the C-C Motif Chemokine Ligand 21.(18–21) In a mouse sepsis model, binding of PDPN to the CLEC-2 on inflammatory macrophages and platelets regulates the immune response.(22) Many studies have reported that PDPN expression in cancer cells is associated with tumor metastasis and malignancy.(23–27) PDPN is expressed not only by cancer cells, but also by cells in the cancer microenvironment, such as cancer-associated fibroblasts, inflammatory macrophages, and T helper 17 cells, and contributes to tumor progression.(28,29) PDPN has also attracted attention as a target for cancer treatment.(28,30) The application of anti-PDPN mAbs in chimeric antigen receptor-T therapies and the development of cancer-specific mAbs have been proposed.(31,32)
We have established different anti-PDPN mAbs for many species by employing a cell-based immunization and screening (CBIS) method, and characterized the targeted epitopes.(33–47) Recently, we developed a novel anti-PDPN mAb, clone PMab-269 (mouse IgG1, kappa), which recognizes the California sea lion (Zalophus californianus) PDPN (seaPDPN).(48) The PMab-269 clearly detected seaPDPN protein in flow cytometry and Western blotting analysis. Furthermore, PMab-269 identified seaPDPN-positive cells in formalin-fixed paraffin-embedded California sea lion specimens, including lung type I alveolar cells, kidney podocytes, and lymphatic endothelial cells in the colon. In this study, we clarified the PMab-269 epitope of seaPDPN using enzyme-linked immunosorbent assay (ELISA).
Materials and Methods
Peptides
The seaPDPN (Accession No.: XP_027432487) peptides, including 10 deletion mutants (Table 1) and 20 point mutants (Table 2), were synthesized by utilizing PEPScreen (Sigma-Aldrich Corp., St. Louis, MO).
Identification of PMab-269 Epitope Using Deletion Mutants
+++, OD655 ≧0.3; −, OD655 < 0.1.
Identification of PMab-269 Epitope Using Point Mutants
+++, OD655 ≧0.3; ++, 0.2≦ OD655 < 0.3; −, OD655 < 0.1.
Enzyme-linked immunosorbent assay
Peptides were immobilized on Nunc Maxisorp 96-well immunoplates (Thermo Fisher Scientific, Inc., Waltham, MA) at a concentration of 10 μg/mL for 30 minutes at 37°C. After washing with phosphate-buffered saline containing 0.05% Tween20 (PBST), wells were blocked with 1% bovine serum albumin-containing PBST for 30 minutes at 37°C. The plates were then incubated with PMab-269 (5 μg/mL),(48) followed by a 1:2000 dilution of peroxidase-conjugated anti-mouse immunoglobulins (Agilent Technologies, Inc., Santa Clara, CA). Enzymatic reactions were performed using the ELISA POD Substrate TMB Kit (Nacalai Tesque, Inc., Kyoto, Japan). Optical density was measured at 655 nm using an iMark microplate reader (Bio-Rad Laboratories, Inc., Berkeley, CA).
Results and Discussion
Epitope mapping using deletion mutants
In our previous study, we developed an anti-seaPDPN mAb (clone PMab-269) using the CBIS method.(48) PMab-269 is suitable for in vitro applications including flow cytometry, Western blotting, and immunohistochemistry. To better characterize the epitope binding site of seaPDPN antigens recognized by PMab-269, we synthesized 10 peptides comprising 23–42 amino acids (aa), 33–52 aa, 43–62 aa, 53–72 aa, 63–82 aa, 73–92 aa, 83–102 aa, 93–112 aa, 103–122 aa, and 113–128 aa, respectively, of the seaPDPN protein sequence (Table 1), and performed ELISA. The results demonstrated that PMab-269 reacted with the 63–82 aa sequence (63-LTALVPINTESVTDFHLEDG-82) of seaPDPN (Fig. 1A). These results are summarized in Figure 1B.

Determination of the PMab-269 epitope of seaPDPN by ELISA using deletion mutants.
Epitope mapping using point mutants
Next, we synthesized 20 different point mutants using the 63–82 aa peptide of seaPDPN (Table 2). The ELISA results demonstrated that PMab-269 reacted with point mutants, such as L63A, T64A, A65G, L66A, V67A, I69A, N70A, T71A, E72A, S73A, V74A, T75A, and G82A, as well as the 63–82 aa wild-type sequence (positive control) (Fig. 2A). In contrast, PMab-269 did not bind to point mutants, such as the P68A, D76A, F77A, H78A, L79A, E80A, and D81A (Fig. 2A), indicating that Pro68, Asp76, Phe77, His78, Leu79, Glu80, and Asp81 are the critical amino acids, which are included in the PMab-269 epitope. As summarized in Figure 2B, the PMab-269 binds to Glu80 and Asp81, which are residues comprising the PLD region. Therefore, PMab-269 might possess neutralizing activity for seaPDPN-induced platelet aggregation.
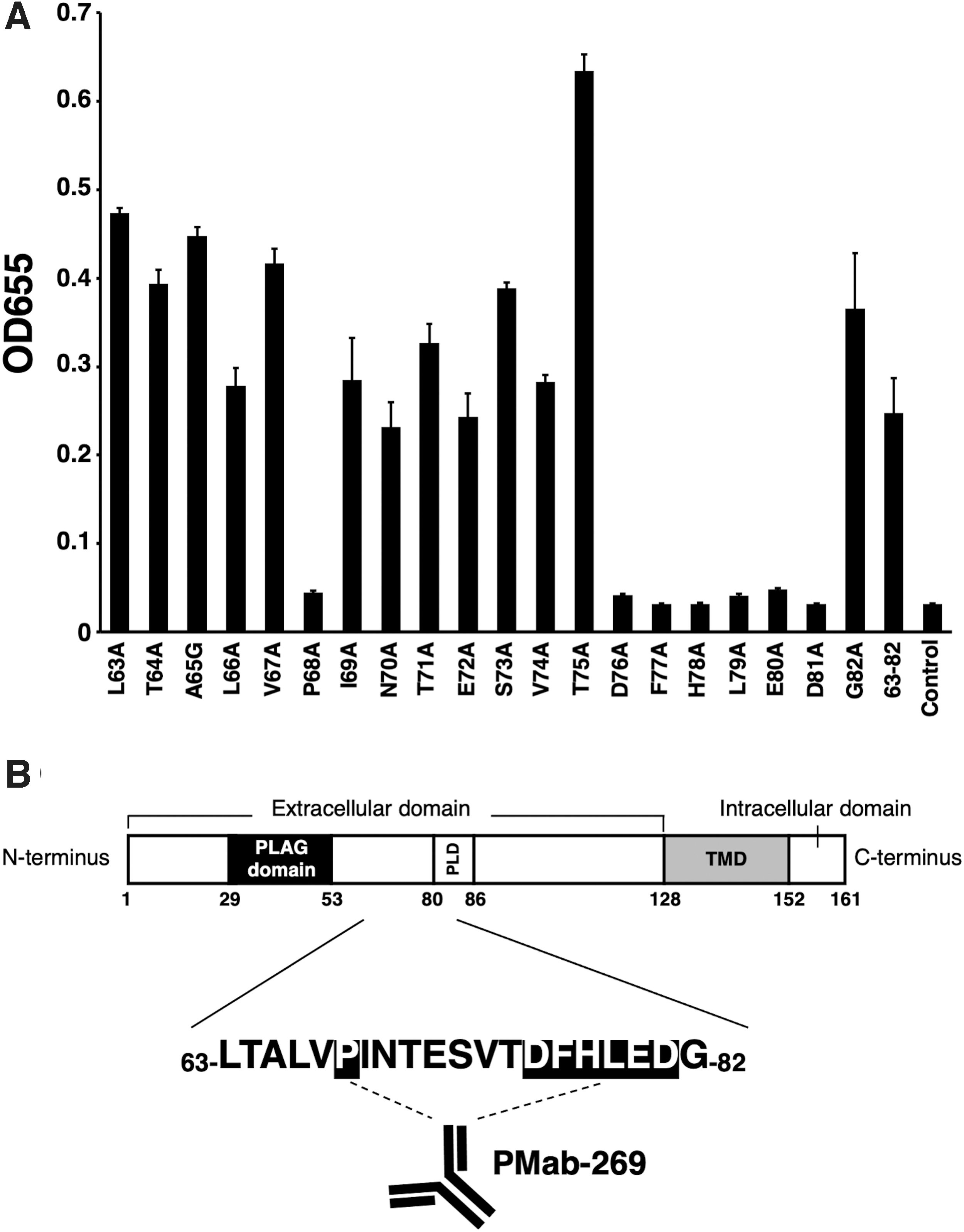
Determination of the PMab-269 epitope of seaPDPN by ELISA using point mutants.
In summary, this study identified the seven amino acids (Pro68, Asp76, Phe77, His78, Leu79, Glu80, and Asp81) as the critical epitope targeted by PMab-269. In the future, the PMab-269 might contribute to pathophysiological studies using the California sea lion model.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported, in part, by Japan Agency for Medical Research and Development (AMED) under Grant Nos. JP21am0401013 (Y.K.) and JP21am0101078 (Y.K.), and by the Japan Society for the Promotion of Science (JSPS) Grants-in-Aid for Scientific Research (KAKENHI) Grant Nos. 21K15523 (to T.A.), 21K07168 (to M.K.K.), 20K16322 (to M.S.), and 19K07705 (to Y.K.).