
Abstract
Rabies is a highly neurotropic disease caused by rabies lyssavirus (RABV). Human rabies vaccines exist for pre- and postexposure prophylaxis; however, after clinical symptoms appear, the disease has an ∼100% mortality rate with no effective treatments available. In our previous study, mouse neuroblastoma cells transfected with a plasmid coding one clone of a single-chain variable fragment (scFv), scFv-P19, against RABV phosphoprotein (RABV-P) derived from an scFv phage-display library, before infection, exhibited reduced viral propagation after infection with the RABV-fixed strain, CVS11. In this study, we conducted epitope mapping of scFv-P19 through indirect fluorescent assay and Western blotting analysis against full-length and N- or C-terminal truncated RABV-P. Our results suggest that scFv-P19 targets a portion containing amino acids 47–52 at the N-terminus, which partially overlaps with the N-terminal nuclear export sequences. This provides insights into the underlying mechanism associated with inhibition of RABV by scFv-P19, while allowing for the design of additional scFv-based therapeutic studies for RABV by integrating appropriate delivery and application systems. Furthermore, the results of this study suggest that scFv-P19 may serve as an effective tool for investigating nuclear trafficking of RABV-P to explore the roles of RABV-P isoforms in rabies pathogenesis.
Introduction
Rabies lyssavirus (RABV), a member of the genus Lyssavirus and family Rhabdoviridae, is highly neurotropic and causes acute infection of the central nervous system in both humans and animals. It is transmitted by the bites or scratches of rabid animals. From the point of entry, RABV travels retrogradely along nerves to the central nervous system. After viral multiplication in the brain causing acute encephalitis, RABV spreads centrifugally and through cell-to-cell transmission to various peripheral tissues, leading to multiorgan failure. The onset of rabies can be prevented by prompt postexposure prophylaxis. However, if clinical symptoms develop, the mortality rate is ∼100%, with no currently available effective treatment options.
We previously investigated whether intracellularly expressed antibodies, intrabodies, against RABV phosphoprotein (RABV-P), could inhibit RABV propagation in neuronal cells.(1) For the intrabodies, we employed a single-chain variable fragment (scFv), comprising the VH and VL regions of the variable antigen-binding site of an immunoglobulin, connected by a short linker sequence.(2) RABV-P was selected as a target because it interacts with various viral or cellular proteins to exert multiple essential functions in viral propagation, including encapsidation of viral genome RNA,(3) acting as a cofactor for the RABV large protein(4) (RABV-L, a viral RNA polymerase), and inhibition of the cellular antiviral system induced by interferons (IFNs).(5)
One of four genetically independent scFv clones derived from an scFv phage-display library, scFv-P19, colocalized with RABV-P in the cytoplasm.(1) Furthermore, scFv-P19, when expressed transiently in mouse neuronal cell lines before RABV infection, severely inhibited RABV propagation and secretion, suggesting that the scFv-based antiviral approach may serve as a candidate for future RABV therapeutic studies if combined with appropriate delivery and application systems; however, more in-depth mechanistic studies are warranted to first elucidate how scFv-P19 elicits its antiviral effects.
In this study, to obtain fundamental information for elucidating the mechanism of RABV inhibition by scFv-P19, we conducted epitope mapping of scFv-P19.
Materials and Methods
Single-chain variable fragment
The selection, expression, and purification of scFv-P19 were performed as previously described.(1) Human single-fold scFv libraries I + J (Tomlinson I+J; a kind gift from MRC Center for Protein Engineering, Cambridge, UK) were used to select phages expressing scFv with the ability to bind either native or denatured RABV-P derived from the fixed RABV strain CVS-11. In the libraries, the scFv sequence was flanked with His-tag- and myc tag-coding sequences and cloned into pIT2 phagemid vectors to express His/myc-tagged scFv.
Immunofluorescence analysis
Plasmid DNA expressing full-length,(6) N-, and C-terminal-truncated RABV-P were generated as described in the Supplementary Data S1. To confirm the expression of the truncated RABV-P, each construct was transfected into HEK293T cells derived from human embryonic kidney cells grown in 96-well plates using the Fugene HD transfection reagent (Promega, WI), following the manufacturer's protocol. Two days after transfection, the expression of each RABV-P variant was confirmed using immunofluorescence analysis (IFA). Details are provided in Supplementary Data S1. The binding ability of scFv-P19 against each RABV-P variant was also confirmed following the same IFA protocol.
Western blotting analysis
HEK293T cells were transfected with RABV-P cDNA as mentioned earlier. Two days after transfection, the cells were lysed with 350 μL of lysis buffer (1% [v/v] Triton-X 100 in PBS[−]) supplemented with one tablet per 10 mL of protease inhibitors (Complete Mini EDTA-free [Sigma-Aldrich]) for 5 minutes on ice. The cell lysates (270 μL) were mixed with 145 μL of loading buffer, including 4 × NuPAGE LDS Sample buffer (Invitrogen) and 10 × reducing agent (Invitrogen) and were sonicated until no longer viscous. Using the remaining cell lysates, protein concentrations were measured using a bicinchoninic acid (BCA) Protein Assay Kit (TaKaRa, Tokyo, Japan).
A total of 5 μg of protein from the cell lysates was subjected to SDS-PAGE using NuPAGE Novex Bis-Tris Mini Gels (Invitrogen) and MES Buffer (Invitrogen). After SDS-PAGE, the proteins were transferred onto PVDF membranes using a semidry transblotter (Bio-Rad, Hercules, CA). The membranes were blocked for 1 hour at room temperature (or overnight at 4°C) with Blocking One reagent (Nakarai, Kyoto, Japan). The detailed protocols for antibody staining, signal development, and image acquisition are provided in the Supplementary Data S1.
Genetic analysis
Amino acid sequences of RABV-P in a total of 566 RABV strains, for which the complete viral genome sequences were registered in GenBank, were aligned and further analyzed using MEGA X software (downloaded from
Results and Discussion
In this study, we conducted epitope mapping of scFv-P19 to elucidate the mechanism of RABV inhibition by scFv-P19. For epitope mapping, full-length RABV-P (P1), N-terminal-truncated RABV-P isoforms, and C-terminal-truncated RABV-P flanked with FLAG/Strept tags (Fig. 1) were transiently expressed in HEK293T cells and analyzed using IFA and Western blotting (WB). The expression of all RABV-P variants was confirmed using anti-FLAG rabbit polyclonal antibody through IFA (data not shown) and WB (Supplementary Fig. S1). Subsequently, the reactivity of scFv-P19 against each variant was examined using IFA (Fig. 2A) and WB (Supplementary Fig. S1A). IFA showed that scFv-P19 reacted positively with P1, P2, and all C-terminal-truncated RABV-P variants (Fig. 2A).
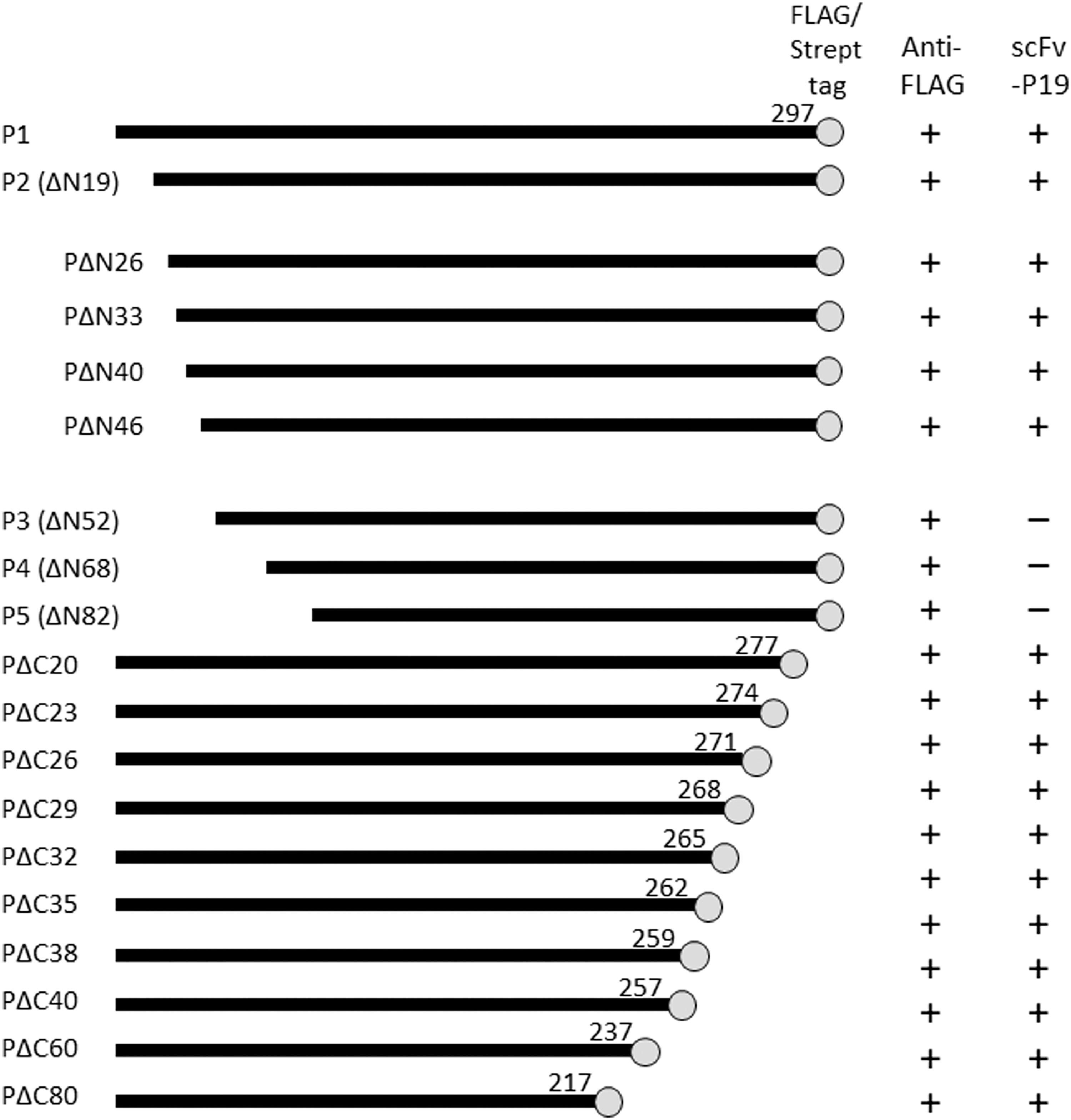
Schematic representation of the full-length and truncated RABV-P used in this study. Black bars represent full-length and truncated RABV-P with respective C-terminal amino acid positions. Gray circles indicate FLAG/Strept tags fused with proteins. The binding of each RABV-P variant with anti-FLAG antibody and scFv-P19 are shown as reactive (+) or nonreactive (–). RABV-P, rabies lyssavirus phosphoprotein; scFv, single-chain variable fragment.
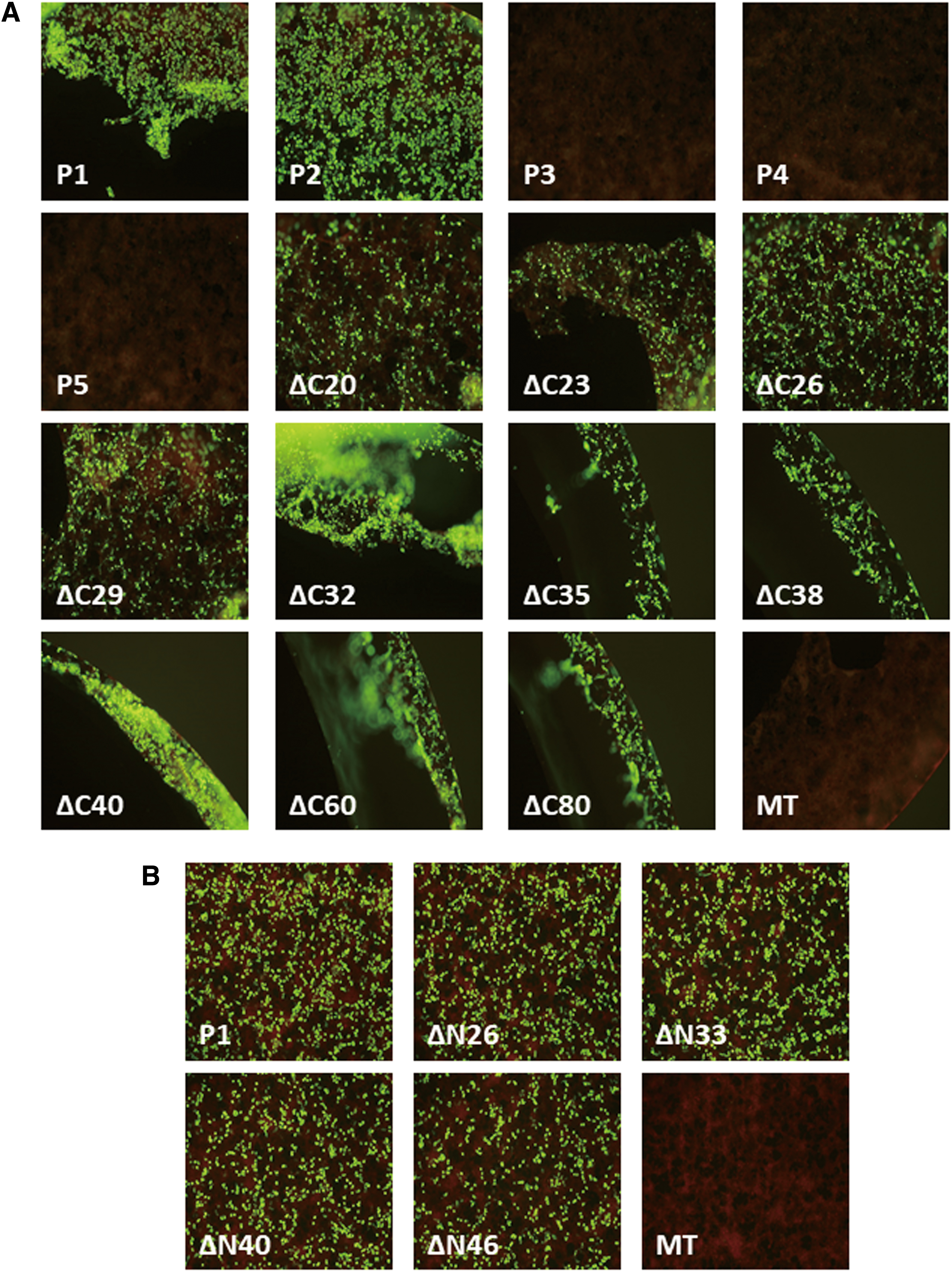
The detection of full-length and truncated RABV-P using IFA. HEK293T cells transiently expressing full-length/truncated RABV-P- and mock-transfected HEK293T cells subjected to IFA for epitope mapping of scFv-P19.
WB also demonstrated that scFv-P19 recognized P1, P2, and all C-terminal-truncated RABV-P variants (Supplementary Fig. S1A), indicating that the scFv-P19 epitope was located between P2 and P3 initiation codons (amino acids [aa] 19–54). To narrow down the range of the scFv-P19 epitope, four additional N-terminal-truncated RABV-P variants, ΔN26, ΔN33, ΔN40, and ΔN46, flanked with FLAG-/Strept-tag (Fig. 1), were transiently expressed in HEK293T cells and analyzed similarly. IFA (Fig. 2B) and WB (Supplementary Fig. S1B) showed that scFv-P19 reacted with ΔN26, ΔN33, ΔN40, and ΔN46, suggesting that the epitope of scFv-P19 exists in the region, including the aa residues 47–52.
To exert multiple functions of RABV-P, five isoforms (P1–P5) interact with various viral and cellular proteins in infected cells. The function of each isoform is associated with the position of the binding domain with other viral/cellular proteins and its intracellular localization.(7) P1, which contains an L-binding domain, N-terminal residues 1–19, acts as an essential cofactor for RABV-L to complete RABV genome transcription and replication.(4) The shorter isoforms (P2–P5) lacking the L-binding domain are involved in other functions, including antagonism of the host cell IFN-dependent immune response.(5)
P1 and P2 are localized primarily in the cytoplasm, whereas P3–P5 are expressed in the nucleus and cytoplasm. Nucleocytoplasmic localization is dominated by the interaction of distinct nuclear trafficking signals,(7) nuclear localization sequence/nuclear export sequences (NES) within each isoform, and cellular proteins involved in nuclear import/export. The cytoplasmic localization of P1 and P2 is thought to be based on the N-terminal NES located at aa residues 49–58, which is found only in P1 and P2. A recent study revealed experimental evidence that RABV-P isoforms play a critical role in rabies pathogenesis.(8)
In this study, we revealed that the scFv-P19 epitope partially overlapped with the N-terminal NES. This is consistent with our previous IFA finding, which revealed that scFv-P19 is colocalized with RABV-P in the cytoplasm.(1) Moreover, in this study we observed that aa residues 47–52, and the adjacent N-terminal NES, were relatively well conserved in RABV-P, which is the least conserved among all RABV proteins (Supplementary Fig. S2).
Our results also suggest that scFv-P19 might affect the nuclear trafficking of isoforms P1 and P2 leading to RABV inhibition. To explore this possibility, a detailed investigation of the intracellular localization of RABV-P isoforms with/without of scFv-P19, as well as quantitative analysis of IFN signaling, are required. Considering that intrabodies flanked with signaling sequences (i.e., ER retention signal) have previously been used to alter the intra- and extracellular localization of target proteins,(9) scFv-P19 with certain modifications might provide potential therapeutic applications in future studies.
In addition, scFv-P19 can be an effective tool for investigating the nuclear trafficking of RABV-P. Considering that recent studies(10,11) suggest that RABV-P may interact with more cellular proteins than previously reported, some of these interactions might be disrupted by scFv-P19 in future studies.
In conclusion, scFv-P19 targets aa 47–52 at the N-terminus, which partially overlaps with the N-terminal NES of RABV-P isoforms, P1 and P2. Thus, scFv-P19 may serve as an effective tool not only for scFv-based therapeutic studies against RABV, but also for investigating the roles of RABV-P isoforms in rabies pathogenesis.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.