
Abstract
The CXC chemokine receptor 6 (CXCR6) is a member of the G protein-coupled receptor family that is highly expressed in helper T type 1 cells, natural killer cells, cytotoxic T lymphocytes, and various type of cells in tumor microenvironment (TME). CXCR6 has been proposed as a therapeutic target against tumors through regulation of the tumor TME. In this study, we developed specific and sensitive monoclonal antibodies (mAbs) for mouse CXCR6 (mCXCR6), which are useful for flow cytometry and Western blotting by N-terminal peptide immunization into rat. The established anti-mCXCR6 mAb, Cx6Mab-1 (rat IgG1, kappa), reacted with not only mCXCR6-overexpressed Chinese hamster ovary-K1 (CHO/mCXCR6) but also mCXCR6-endogenously expressed cell lines, such as P388 (mouse lymphoid neoplasm) and J774-1 (mouse macrophage-like) through flow cytometry. Kinetic analyses using flow cytometry indicated that the dissociation constants (KD) of Cx6Mab-1 for CHO/mCXCR6, P388, and J774-1 cells were 1.7 × 10−9 M, 3.4 × 10−7 M, and 3.8 × 10−7 M, respectively. Furthermore, Cx6Mab-1 could detect endogenous mCXCR6 in P388 and J774-1 cells by Western blotting. These results indicated that Cx6Mab-1 is useful for detecting mCXCR6 by flow cytometry and Western blotting, and provides a possibility for targeting CXCR6-expressing cells in vivo experiments.
Introduction
The CXC chemokine receptor 6 (CXCR6) is a chemokine receptor for CXCL16. CXCR6 is primarily expressed in T cells, including cytotoxic T lymphocytes (CTLs), helper T type 1 (Th1) cells, natural killer (NK) cells, and NK T cells. Upon the chemokines binding to CXCR6, the chemokines can stimulate intracellular signaling pathway in CXCR6-expressing cells, which mediate various cellular functions, including the infiltration into target tissues.(1) CXCR6 has been reported to play a critical role in lymphocyte homeostasis in the liver, based on the constitutive secretion of CXCL16 in liver sinusoids. CXCL16 supports the maintenance of CXCR6-expressing NK, NK T, and CD8+ memory T cells in liver.(2)
CXCL16 is the largest ligand compared with the other CXC chemokines. CXCL16 has been found in soluble form (sCXCL16) and transmembrane form (TM-CXCL16).(3) Therefore, the forward and reverse signaling pathways are transduced through TM-CXCL16 and CXCR6 interaction. In forward signaling, sCXCL16 or TM-CXCL16 binds to CXCR6 that promotes signal transduction from CXCR6. In reverse signaling, CXCR6-bound TM-CXCL16 mediates the signal transduction.(4)
The expression of CXCL16 and CXCR6 has been observed in numerous human cancers, including pancreatic cancer, thyroid carcinoma, and breast cancer, and influences both tumor and the tumor microenvironment (TME).(4) The growing number of evidence has shown the relationship between chronic inflammation and tumor development.(5) CXCL16–CXCR6 axis plays critical roles in not only lymphocyte recruitment to tumor sites, but also in tumor development through activation of tumor invasiveness and cancer-associated fibroblasts (CAFs).(4)
Using the Cell-Based Immunization and Screening (CBIS) method, we have developed many monoclonal antibodies (mAbs) against membrane proteins, such as anti-CD19,(6) anti-CD20,(7,8) anti-CD44,(9) anti-CD133,(10) anti-EpCAM,(11,12) anti-HER3,(13) anti-KLRG1,(14) anti-TIGIT,(15) anti-TROP2,(16,17) anti-programmed cell death ligand 1 (PD-L1),(18) and anti-podoplanin(19–30) mAbs. The CBIS method includes the immunization of antigen-overexpressing cells and the high-throughput hybridoma screening using flow cytometry.
Anti-chemokine receptor mAbs, including anti-mouse CCR3,(31) anti-mouse CCR8,(32) and anti-human CCR9(33) mAbs, were also successfully developed using the CBIS method. Furthermore, we have developed several anti-chemokine receptor mAbs by peptide immunization.(34) This study developed anti-mouse CXCR6 (mCXCR6) mAbs using N-terminal peptide immunization.
Materials and Methods
Preparation of cell lines
Chinese hamster ovary (CHO)-K1 and P3X63Ag8U.1 (P3U1) cells were obtained from the American Type Culture Collection (Manassas, VA). P388 (mouse lymphoid neoplasm) and J774-1 (mouse macrophage-like) cells were obtained from the Cell Resource Center for Biomedical Research Institute of Development, Aging and Cancer Tohoku University (Miyagi, Japan). The synthesized DNA (Eurofins Genomics KK, Tokyo, Japan) encoding mCXCR6 (Accession No. NM_030712) was subsequently subcloned into a pCAG-Ble vector (FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan).
Then, the mCXCR6 plasmid was transfected into CHO-K1 cells, using a neon transfection system (Thermo Fisher Scientific, Inc., Waltham, MA). Stable transfectants were established through cell sorting using a cell sorter (SH800; Sony Corp., Tokyo, Japan), after which cultivation in a medium, containing 0.5 mg/mL of Zeocin (InvivoGen, San Diego, CA), was conducted.
CHO-K1, mCXCR6-overexpressed CHO-K1 (CHO/mCXCR6), P3U1, P388, and J774-1 were cultured in a Roswell Park Memorial Institute (RPMI) 1640 medium (Nacalai Tesque, Inc., Kyoto, Japan), with 10% heat-inactivated fetal bovine serum (FBS; Thermo Fisher Scientific, Inc.), 100 units/mL of penicillin, 100 μg/mL of streptomycin, and 0.25 μg/mL of amphotericin B (Nacalai Tesque, Inc.). Cells were grown in a humidified incubator at 37°C, at an atmosphere of 5% CO2 and 95% air.
Antibodies
An anti-mCXCR6 mAb (clone SA051D1) was purchased from BioLegend (San Diego, CA). A secondary Alexa Fluor 488-conjugated anti-rat IgG was purchased from Cell Signaling Technology, Inc. (Danvers, MA).
Peptides
Eurofins Genomics KK (Tokyo, Japan) synthesized a partial sequence of the N-terminal extracellular region of mCXCR6 (1-MDDGHQESALYDGHYEGDF-19) plus C-terminal cysteine. Subsequently, the keyhole limpet hemocyanin (KLH) was conjugated at the C-terminus of the peptide.
Production of hybridomas
A Sprague–Dawley rat (5 weeks old) were purchased from CLEA Japan (Tokyo, Japan). Animals were housed under specific pathogen-free conditions. All animal experiments were also conducted according to relevant guidelines and regulations to minimize animal suffering and distress in the laboratory. The Animal Care and Use Committee of Tohoku University (Permit No. 2019NiA-001) approved animal experiments. Rats were monitored daily for health during the full 4-week duration of the experiment. A reduction of >25% of the total body weight was defined as a humane endpoint. During sacrifice, the rat was euthanized through cervical dislocation, after which death was verified through respiratory and cardiac arrest.
To develop mAbs against mCXCR6, we intraperitoneally immunized one rat with 100 μg of the KLH-conjugated mCXCR6 peptide plus Imject Alum (Thermo Fisher Scientific, Inc.). The procedure included three additional immunization every week (100 μg/rat), which was followed by a final booster intraperitoneal injection (100 μg/rat), 2 days before the harvest of spleen cells. Harvested spleen cells were subsequently fused with P3U1 cells, using PEG1500 (Roche Diagnostics, Indianapolis, IN), after which hybridomas were grown in an RPMI medium with 10% FBS, 100 units/mL of penicillin, 100 μg/mL of streptomycin, and 0.25 μg/mL of amphotericin B.
For the hybridoma selection, hypoxanthine, aminopterin, and thymidine (HAT; Thermo Fisher Scientific, Inc.) were added into the medium. Supernatants were subsequently screened using enzyme-linked immunosorbent assay (ELISA) with the mCXCR6 peptide, followed by flow cytometry, using CHO/mCXCR6, CHO-K1, P388, and J774-1 cells.
Enzyme-linked immunosorbent assay
The synthesized peptide (MDDGHQESALYDGHYEGDFC) was immobilized on Nunc Maxisorp 96-well immunoplates (Thermo Fisher Scientific, Inc.) at a concentration of 1 μg/mL for 30 minutes at 37°C. After washing with phosphate-buffered saline (PBS) containing 0.05% Tween 20 (PBST; Nacalai Tesque, Inc.), wells were blocked with 1% bovine serum albumin (BSA)-containing PBST for 30 minutes at 37°C. Plates were then incubated with supernatants of hybridomas, followed by peroxidase-conjugated anti-rat immunoglobulins (1:20,000 diluted; Sigma-Aldrich Corp., St. Louis, MO). Next, enzymatic reactions were conducted, using ELISA POD Substrate TMB Kit (Nacalai Tesque, Inc.), followed by measurement of the optical density at 655 nm, using iMark microplate reader (Bio-Rad Laboratories, Inc., Berkeley, CA).
Purification of mAbs
The cultured supernatants of Cx6Mab-1–producing hybridomas were collected through centrifugation at 2330 × g for 5 minutes, followed by filtration using Steritop (0.22 μm; Merck KGaA, Darmstadt, Germany). Filtered supernatants were subsequently applied to 1 mL of Ab-Capcher ExTra (ProteNova Co., Ltd., Kagawa, Japan). After washing with PBS, bound antibodies were eluted with IgG Elution Buffer (Cat. No.: 21009, Thermo Fisher Scientific, Inc.), followed by immediate neutralization of eluates, using 1 M Tris-HCl (pH 8.0). Finally, eluates were concentrated, after which PBS was used to replace with the elution buffer, using Amicon Ultra (Merck KGaA).
Flow cytometric analyses
CHO-K1 and CHO/mCXCR6 were harvested after a brief exposure to 0.25% trypsin and 1 mM ethylenediaminetetraacetic acid (EDTA; Nacalai Tesque, Inc.). Cells were subsequently washed with 0.1% BSA in PBS and treated with 0.01, 0.1, 1, and 10 μg/mL primary mAbs for 30 minutes at 4°C. Then, cells were treated with Alexa Fluor 488-conjugated anti-rat IgG (1:2000; Cell Signaling Technology, Inc.), after which fluorescence data were collected using the SA3800 Cell Analyzer (Sony Biotechnology Corp.).
Determination of dissociation constant (KD) through flow cytometry
CHO/mCXCR6, P388, and J774-1 were suspended in 100 μL serially diluted anti-mCXCR6 mAbs, after which 50 μL Alexa Fluor 488-conjugated anti-rat IgG (1:200; Cell Signaling Technology, Inc.) was added. Then, fluorescence data were collected, using BD FACSLyric (BD Biosciences, Franklin Lakes, NJ). The KD was subsequently calculated by fitting saturation binding curves to the built-in one-site binding models in GraphPad PRISM 8 (GraphPad Software, Inc., La Jolla, CA).
Western blot analysis
Cell lysates were prepared by hypotonic cell lysis buffer (20 mM HEPES–KOH, pH 7.5, 10 mM KCl, 1.5 mM MgCl2, 0.2 mM EDTA, 0.1% Triton-X100), and were boiled in sodium dodecyl sulfate (SDS) sample buffer (Nacalai Tesque, Inc.). Proteins (10 μg for CHO-K1 and CHO/mCXCR6 or 15 μg for P388 and J774-1) were separated on 5%–20% polyacrylamide gels (FUJIFILM Wako Pure Chemical Corporation) and transferred onto polyvinylidene difluoride (PVDF) membranes (Merck KGaA).
After blocking with 4% skim milk (Nacalai Tesque, Inc.) in PBST, membranes were incubated with 10 μg/mL of Cx6Mab-1, 1 μg/mL of an anti-isocitrate dehydrogenase 1 (IDH1; RcMab-1)(35,36) or 1 μg/mL of an anti-β-actin mAb (clone AC-15; Sigma-Aldrich Corp.). Membranes were then incubated with either peroxidase-conjugated anti-rat and anti-mouse immunoglobulins (diluted 1:1000; Agilent Technologies, Inc., Santa Clara, CA). Finally, protein bands were detected with ImmunoStar LD (FUJIFILM Wako Pure Chemical Corporation) using a Sayaca-Imager (DRC Co. Ltd., Tokyo, Japan).
Results
Development of anti-mCXCR6 mAbs by peptide immunization
To develop anti-mCXCR6 mAbs, one rat was immunized with the KLH-conjugated mCXCR6 peptide (Fig. 1A). Spleen was then excised from the rat, after which splenocytes were fused with myeloma P3U1 cells (Fig. 1B). Developed hybridomas were subsequently seeded into twenty 96-well plates and cultivated for 6 days. Then, positive wells for the naked mCXCR6 peptide were selected using ELISA, followed by the selection of mCXCR6-expressing cell-reactive and CHO-K1 nonreactive supernatants, using flow cytometry (Fig. 1C). The ELISA screening identified 26 out of 1914 wells (1.36%), which strongly reacted with the naked mCXCR6 peptide.

A schematic illustration showing the production of anti-mCXCR6 mAbs.
The flow cytometric screening identified 12 out of the 26 wells (46.2%), which exhibited strong signals to CHO/mCXCR6 cells, but not to CHO-K1 cells. In addition, positive signals were also obtained in endogenous mCXCR6-expressing P388 and J774-1 cells in 5 out of the 12 wells (41.7%). After the limiting dilution and several additional screenings, an anti-mCXCR6 mAb (clone Cx6Mab-1; rat IgG1, kappa) was finally established (Fig. 1D).
Flow cytometric analysis
We conducted flow cytometry using two anti-mCXCR6 mAbs: Cx6Mab-1 and SA051D1 against CHO/mCXCR6, P388, and J774-1 cell lines. Both Cx6Mab-1 and SA051D1 recognized CHO/mCXCR6 cells dose-dependently at 10, 1, and 0.1 μg/mL. Cx6Mab-1 could recognize CHO/mCXCR6 cells at low concentration of 0.01 μg/mL; in contrast, SA051D1 needed >0.1 μg/mL for the detection of CHO/mCXCR6 (Fig. 2A). Parental CHO-K1 cells were not recognized even at 10 μg/mL of both mAbs (Fig. 2B). Cx6Mab-1 and SA051D1 reacted with P388 and J774-1 cells at >1 μg/mL (Fig. 3A, B). These results suggested that Cx6Mab-1 specifically recognizes mCXCR6, and is also useful for detecting endogenous mCXCR6 by flow cytometry.
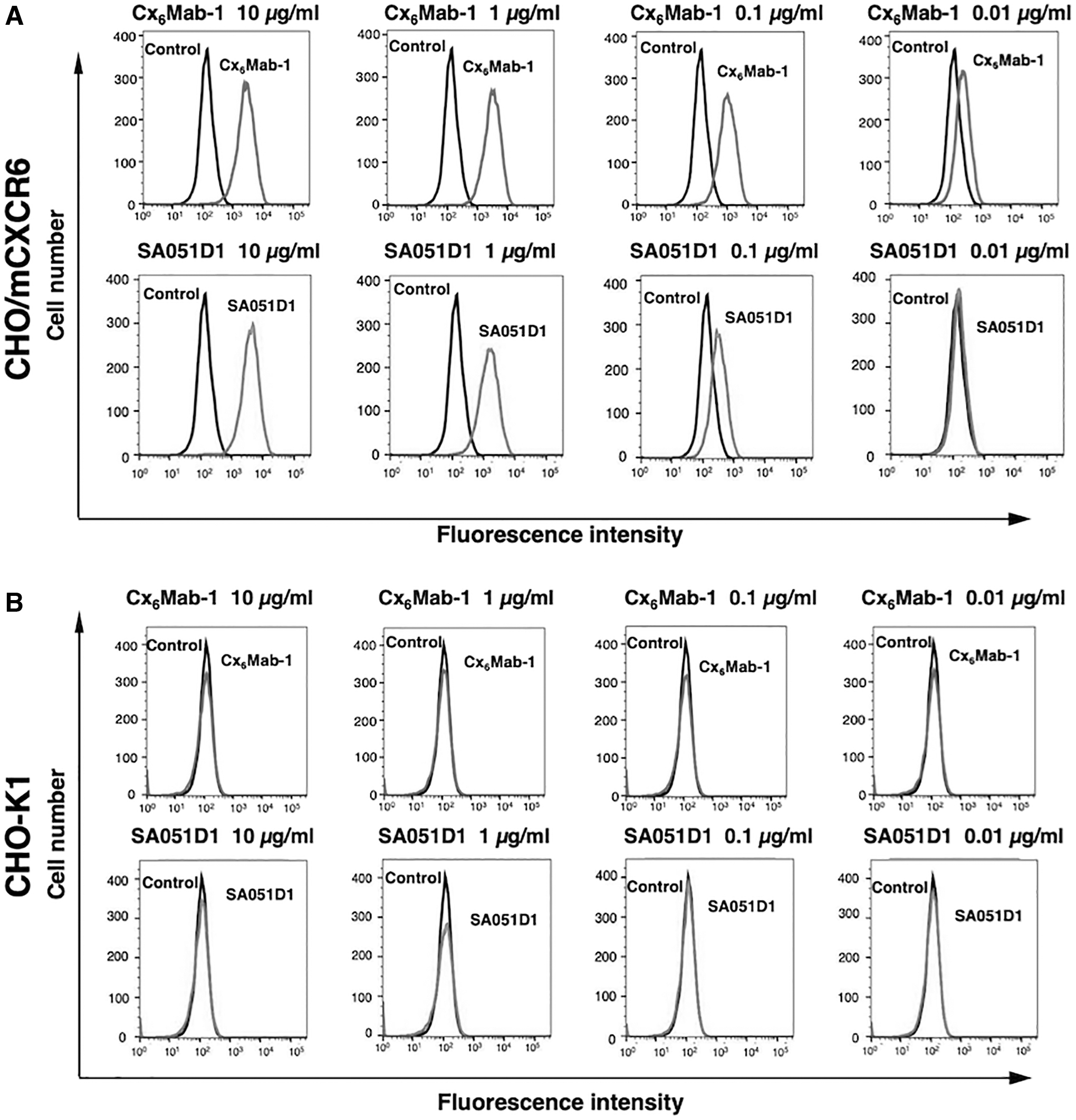
Flow cytometry to mCXCR6 overexpressing cells using anti-mCXCR6 mAbs. CHO/mCXCR6
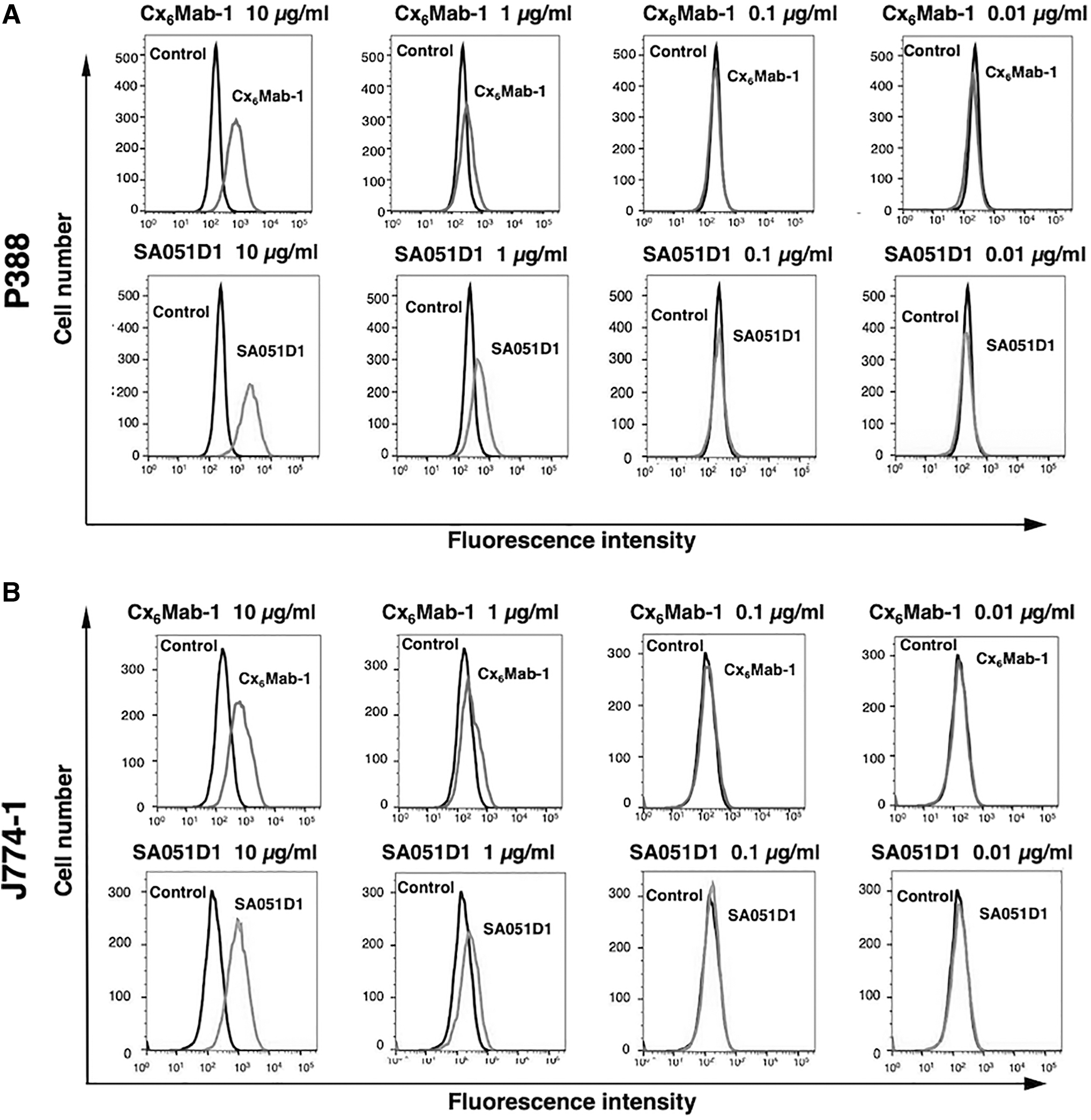
Flow cytometry to endogenously mCXCR6-expressing cells using anti-mCXCR6 mAbs. P388
Kinetic analysis of the interaction between Cx6Mab-1 and mCXCR6-expressing cells through flow cytometry
To determine the KD of Cx6Mab-1 with mCXCR6-expressing cells, we conducted kinetic analysis of the interaction between Cx6Mab-1 and CHO/mCXCR6, P388, and J774-1 cell lines using flow cytometry. The geometric mean of the fluorescence intensity was then plotted versus the concentration of Cx6Mab-1. The KD of Cx6Mab-1 for CHO/mCXCR6, P388, and J774-1 cells was determined as 1.7 × 10−9 M, 3.4 × 10−7 M, and 3.8 × 10−7 M, respectively (Fig. 4). These results indicate that Cx6Mab-1 possesses the high affinity for CHO/mCXCR6 and the moderate affinity for P388 and J774-1 cells.
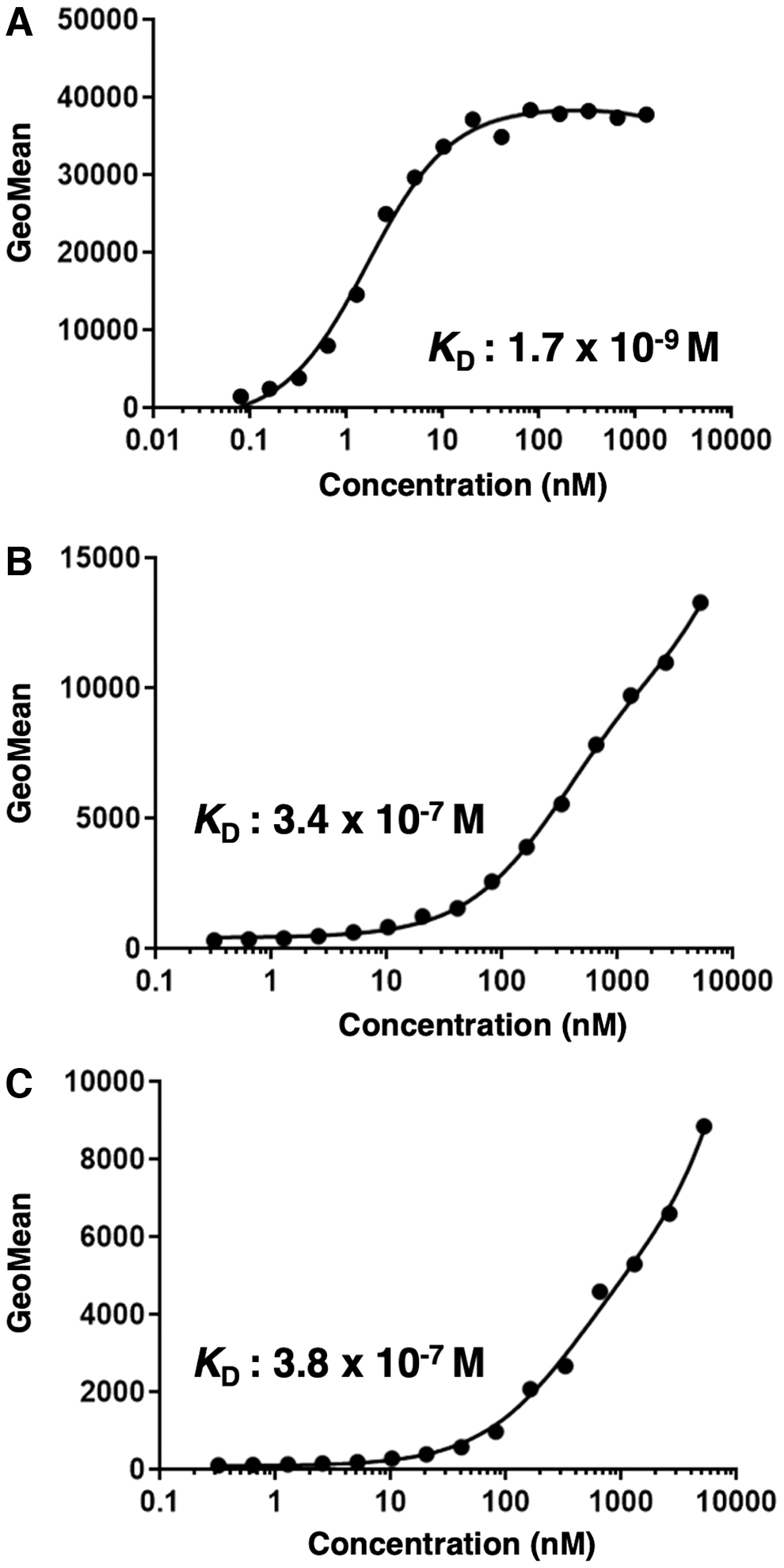
The determination of the binding affinity of Cx6Mab-1. CHO/mCXCR6
Western blot analysis
Western blotting was performed to further assess the sensitivity of Cx6Mab-1. Lysates of CHO-K1 and CHO/mCXCR6 cells were probed. As shown in Figure 5A, Cx6Mab-1 detected mCXCR6 as a ∼40-kDa main band. However, Cx6Mab-1 did not detect any bands from lysates of CHO-K1 cells. Next, we examined the detection of endogenous mCXCR6 using lysates from P388 and J774-1 cells. As shown in Figure 5B, Cx6Mab-1 could detect mCXCR6 as a ∼40-kDa main band, and double bands between 75 and 100 kDa from lysates of P388 and J774-1 cells. Similar molecular weight bands were also detected in lysates of CHO/mCXCR6 cells (Fig. 5A). These results suggest that Cx6Mab-1 can detect both exogenous and endogenous mCXCR6.

Western blotting using Cx6Mab-1.
Discussion
In this study, we developed novel anti-mCXCR6 mAbs using N-terminal peptide immunization methods, and investigated the usefulness for flow cytometry and Western blotting for detecting the endogenous mCXCR6. The synthesized peptide immunization is a simple strategy and possesses several advantages to generate anti-CXCR mAbs since the N-terminal regions of CXCR families are largely and linearly exposed on the cell surface.(37) The neutralizing activity could be expected because the N-terminus of CXCR families is known to be the ligand-binding domain.(37)
Although the three-dimensional structure of CXCR6 and its ligand CXCL16 has not been solved experimentally, a homology model of CXCR6–CXCL16 was reported through coarse-grained molecular dynamics simulations. According to the simulations, the N-terminal region of CXCR6 in complex with CXCL16 is positioned in the middle of extracellular loops, triggers the system's evolution, rotations, movements, and transmembrane interactions, and changes the inactive state to an active state.(38) Since the CXCL16-CXCR6 interaction mediates the forward and reverse signaling pathways,(3,4) further investigation is required to evaluate the effect of Cx6Mab-1 to these pathways.
CXCL16–CXCR6 axis has been reported to possess the complex roles in TME. It mediates tumor progression directly through promoting survival, proliferation, and metastasis of tumor cells or indirectly through enhancing lymphocytes, macrophages, and mesenchymal stem cells (MSCs) infiltration.(4) The signaling of CXCR6 in recruited MSCs leads to the conversion into CAFs.(39) Furthermore, the recruitment of tumor-promoting M2 macrophages is also promoted.(40) These events play critical roles in tumor progression through activating the metastatic programs and angiogenesis.
In our previous studies, we changed the isotype of mAbs into mouse IgG2a to retain antibody-dependent cellular cytotoxicity (ADCC) and complement-dependent cytotoxicity (CDC), which caused high antitumor activities in mice xenograft models.(41–43) Since the subclass of Cx6Mab-1 is rat IgG1, it does not possess ADCC and CDC. Therefore, in further studies, the subclass of Cx6Mab-1 will be converted into mouse IgG2a to evaluate the effect to CXCR6-expressing tumor cells, CAFs, and macrophages in mice tumor models.
CXCR6 mediates the conversion of stem-like memory cells into effector-like CTLs. In TME, CCR7+ dendritic cells exist in perivascular niche, and secrete CXCL16 and trans-present the survival cytokine interleukin-15 (IL-15). CXCR6 expression and IL-15 trans-presentation are critical for the survival and local expansion of effector-like CTLs in TME to potentiate their antitumor response.(44) These observations indicate a critical function of CXCR6 for sustaining antitumor immune responses of CD8+ CTLs.
In tumor immune therapy, the infiltration of CTLs into tumor is important factors to predict the efficacy.(45,46) The PD-1+ effector-like CTLs uniformly express CXCR6 at the highest level.(44) Therefore, the diagnostic use of Cx6Mab-1 to examine the CXCR6 expression in tumor-infiltrating CTLs could provide the important information for combination therapy of immune checkpoint inhibitor with CXCR6 agonists developed in the future.
Footnotes
Author Disclosure Statement
The authors have no conflict of interest.
Funding Information
This research was supported in part by Japan Agency for Medical Research and Development (AMED) under Grant numbers JP22ama121008 (to Y.K.), JP22am0401013 (to Y.K.), JP22bm1004001 (to Y.K.), JP22ck0106730 (to Y.K.), and JP21am0101078 (to Y.K.).